

摘要
此前,对尼安德特人的基因组分析为他们的种群历史以及与现代人的关系提供了洞见1,2,3.,4,5,6,7,8,但尼安德特人社区的社会组织仍然知之甚少。在这里,我们介绍了来自西伯利亚南部阿尔泰山脉两个旧石器时代中期遗址的13个尼安德特人的基因数据:11个来自Chagyrskaya洞穴9,10还有2个来自Okladnikov洞穴11——这是迄今为止对尼安德特人进行的最大规模的基因研究之一。我们使用杂交捕获技术获得全基因组核数据,以及线粒体和y染色体序列。一些Chagyrskaya个体是近亲,包括一对父女和一对二级亲属,这表明至少有一些个体生活在同一时期。这些个体中有多达三分之一的基因组具有长段纯合性,这表明Chagyrskaya尼安德特人是一个小社区的一部分。此外,y染色体的多样性比线粒体的多样性低一个数量级,我们发现这种模式最好地解释了女性在社区之间的迁移。因此,这里提出的基因数据提供了一个与世隔绝的尼安德特人社区的社会组织的详细文件,该社区位于他们已知活动范围的最东端。
主要
约43万年前,尼安德特人占领了欧亚大陆西部8,12直到大约4万年前它们灭绝13.据报道,来自14个考古遗址的18具骸骨的基因组尺度数据1,2,3.,4,5,6,7,8跨越了尼安德特人的历史,跨越了他们已知地理范围的大部分,东至西伯利亚南部的阿尔泰山脉。这些数据对尼安德特人的种群进行了广泛的概述,表明随着时间和空间的推移,存在着多个不同的尼安德特人种群1,2,14.
然而,在这段时间内,欧亚大陆任何地区的尼安德特人社区内部和之间的基因关系和社会组织知之甚少。
所谓“社会组织”,指的是一个社区的规模、性别组成和时空凝聚力15.我们将社区定义为一组可能生活在同一地点的个体,而将“人口”一词保留给在更广泛的地理区域内广泛联系的一组社区。
根据脚印化石16,17以及场地使用的空间模式18在美国,之前对尼安德特人社区社会组织的研究表明,尼安德特人可能生活在小型社区中。此外,来自6个成年尼安德特人的部分线粒体DNA (mtDNA)序列已被用于表明尼安德特人可能是父系的19尽管这一建议一直存在争议20..
在这里,我们利用从南西伯利亚(俄罗斯)两个彼此靠近的地点——chagyrskaya和Okladnikov洞穴中发现的13个个体的遗骸的核、y染色体和mtDNA数据来探索尼安德特人的社会组织1和无花果。1).

一个,所有尼安德特人遗址的位置(有多个尼安德特人的遗址的个体数量在括号中给出),以及西伯利亚南部阿尔泰地区的Chagyrskaya和Okladnikov洞穴的特写。b,核基因组按覆盖范围排序,并按位点进行颜色编码(蓝色,本研究中的Chagyrskaya;橙色,Okladnikov来自这项研究;灰色,以前发表在参考文献。1,2,3.,4,5,6,7,8).c的mtDNA序列的最大似然树,该序列来自本研究中包括的尼安德特人个体,在已知的古人蛋白变异的背景下。参考基因组为rCRS,现代人的登录号为东亚人(AF346973)、欧洲人(AF346981)和非洲人(AF381988)。Okladnikov 2指参考文献中的mtDNA序列。41(此标本在扩展数据表中被列为Okladnikov 141).参考文献中的数据。1,2,3.,4,6,30.,41,42,43,44,45,46,47,48,49.d4个Chagyrskaya个体的Y染色体的6.9 Mb,覆盖不止一个,以及先前发表的3个尼安德特人、2个丹尼索万人和4个现代人的Y染色体数据,基于共识调用的最大似然树。参考基因组为hg19。参考文献中的数据。26,50,51,52,53.在c而且d,单倍群显示为当今人类人口。
考古遗址及遗迹
Chagyrskaya和Okladnikov洞穴,位于阿尔泰山脉的山麓。1和扩展数据图。1而且2),被认为主要用作短期狩猎营地11,21.他们是三个已知的遗址中的两个,在那里发现了一个独特的Sibiryachikha中期旧石器时代工业(第三个是上Sibiryachikha洞穴)9,10,22,23(补充图。1.6).Chagyrskaya和Okladnikov洞穴的Sibiryachikha工业与Denisova洞穴(位于以东约100公里处)的中古旧石器时代工业不同,在那里也发现了尼安德特人的遗骸2.
沉积物的光学测年和野牛骨骼的放射性碳测年表明,Chagyrskaya洞穴中尼安德特人居住的沉积物积累于5.9万至5.1万年前10.我们从两块木炭和一块尼安德特人的骨头(Chagyrskaya 9)中获得了额外的放射性碳年龄,所有这些都比现在早5万年以上(补充表)1.3).这些年代与较短的沉积时期(几千年或更短)相吻合,这与所有尼安德特人地层中存在类似的考古工业是一致的10(扩展数据图。2).
对于Okladnikov洞穴,我们对三个尼安德特人标本(包括Okladnikov 15)使用羟脯氨酸单氨基酸放射性碳年龄来限制尼安德特人占领的时间(表1和扩展数据表1),表明它们至少有44000年的历史(补充表1.4).我们的年龄估计与动物骨骼的铀系列年龄一致,并支持先前的建议,即从胶原蛋白部分获得的较年轻的放射性碳年龄反映了污染物的不完全清除24(补充信息部分1).因此,考古和年代学数据表明,占据这两个地点的尼安德特人可能大致是同时代的。
此前对来自Chagyrskaya洞穴的尼安德特人(Chagyrskaya 8)和更早的来自Denisova洞穴的尼安德特人(Denisova 5,“阿尔泰尼安德特人”)的高覆盖基因组分析显示,他们属于不同的种群5.尼安德特人母亲和丹尼索瓦人父亲的第一代后代(Denisova 11)显示,尼安德特人母亲与Chagyrskaya 8比其他尼安德特人更相似5,25.
数据采集和性别确定
我们从Chagyrskaya洞穴的17个标本和Okladnikov洞穴的10个标本中取样了1-64毫克的牙齿或骨粉。其中,15个来自Chagyrskaya, 2个来自Okladnikov,提取了古代DNA1,扩展数据表1和补充数据1),从中我们总共生成了85个单链DNA文库(补充信息部分)2).所有的库都对mtDNA序列进行了富集(补充信息部分)3.使用新设计的包含643,472个跨基因组横向多态性的核捕获阵列(补充信息部分)对49个文库(选择高序列产量和当今人类污染水平)进行核DNA富集5).在该序列中,271,306个遗址在4个已发表的高覆盖率古人类(3个尼安德特人和1个丹尼索瓦人)中有所不同。2,3.,5,14372166个地点在当今非洲人口中分离出来,或固定在当今人类和古代古人类之间。每个化石的平均核DNA覆盖率在0.04- 12.3倍之间。1 b),而目前人类污染的估计范围为0.1%至3.2%(补充表)5.4).
我们利用X染色体和常染色体之间的覆盖率差异确定了17具遗体的遗传性别(补充图。5.5),并发现6具遗骸来自女性。对于这11具男性遗体,我们对其y染色体序列进行了约6.9兆酶(Mb)的富集26(补充信息部分4),其覆盖率介于0.02- 42.2倍之间(补充表4.3).
识别的亲戚
为了确定是否有任何遗骸来自相关个体,我们从捕获阵列中250,785个位点随机抽样1个等位基因,计算了17具遗骸之间的核DNA差异,这些等位基因在高覆盖率古个体中是可变的(不包括Chagyrskaya 8的特定变体)(补充信息部分)5).对于有亲缘关系的个体来说,这种差异会更低,因为他们从最近的共同祖先那里继承了部分基因组。我们将这个散度归一化(p0)是所有比较中DNA差异的中位数。使用这种方法27,我们可以检测到二级关系;我们认为除此之外的一切都无关紧要。我们预计p0= 1表示遗骸的远亲关系大于二级亲属,p0二级亲属= 0.875,p0一级亲属= 0.75p0同卵双胞胎或同一个体的遗骸= 0.527.我们还研究了线粒体DNA的异质性(也就是说,当个体携带的线粒体的DNA序列不同时)(补充表)3.2)以确定密切的基因关系28.由于异质体可以从母亲传给孩子,通常持续不到三代29,它们出现在不同的遗骸中,表明它们来自同一个人或与母亲关系密切的人。为了区分遗骸(即骨骼和牙齿样本)和个人,我们用数字表示前者,用字母表示后者1).
我们发现了一颗乳牙(Chagyrskaya 19)和两颗恒牙(Chagyrskaya 13和Chagyrskaya 63)。令人惊讶的是,尽管它们的发育阶段不同,遗传数据表明它们属于同一个体(Chagyrskaya G;平均p0= 0.53)3).与此一致的是,所有三颗牙齿都来自一名男性,携带相同的mtdna,包括3961位的异质性,频率相似,为60.7-78.5%(补充表)3.2).几乎完全吸收的乳牙牙根表明它是自然脱落的(补充资料部分)1).根据磨损和牙根发育的模式,我们推断恒牙来自一个9 - 15岁的个体,这名男性可能在乳牙脱落前后死亡。
我们还确定了另外两组拥有多个化石的个体:Chagyrskaya C由Chagyrskaya 6(下颌骨)和Chagyrskaya 14(永久门牙)所代表(补充信息部分)1),这可由形态学拟合、相同的mtDNA序列(包括共享的异质性)和较低的核散度(p0= 0.65;95%置信区间,0.34-0.78)(图1 c,扩展数据图。3和补充表3.2而且7.1).同样,Chagyrskaya F由Chagyrskaya 12和之前测序的Chagyrskaya表示5Chagyrskaya 8 (p0= 0.46;95%置信区间,0.41-0.46)(补充表7.1).
其中一个成年雄性个体Chagyrskaya D与该群体中的其他多个个体密切相关。我们发现他和一名少女Chagyrskaya H有一级关系。p0= 0.77;95%置信区间,0.72-0.82)。一级亲属有三种可能的男女组合:母子、兄妹或父女。然而,由于这两个个体携带不同的线粒体基因组(图。1 c),我们得出结论Chagyrskaya H是Chagyrskaya D的女儿。
此外,它的mtDNA与另外两只雄性Chagyrskaya C和Chagyrskaya E相同3.2),包括545位(G> a)的mtDNA共享异质性,Chagyrskaya D的a频率为42-54%,Chagyrskaya E的a频率为20-41%,Chagyrskaya C的a频率为23-30%。因此,这些人很可能是近母系亲属(例如,他们可能共享一个祖母,因此可能是第四级亲属)。然而,Chagyrskaya C和Chagyrskaya D之间的关系超出了我们方法的解决范围(p0= 1.05;95%置信区间,0.94-1.16)。Chagyrskaya E的覆盖率较低(补充表5.4)和大量人为和非人为污染(补充表)5.3).纠正非人为污染后(补充表7.1),我们将Chagyrskaya E鉴定为Chagyrskaya D的一级亲缘或完全相同(p0= 0.64;95%置信区间,0.48-0.79)。由于我们不能确信Chagyrskaya E是一个不同的个体,我们从进一步的分析中删除了样本。
Chagyrskaya C、D和H之间的密切关系表明它们是同时代的。此外,我们发现Chagyrskaya A(雄性)和L(雌性)是二级亲缘关系(p0= 0.85;95%置信区间,0.77-0.91)。虽然稀疏的数据使我们无法确定确切的关系,但它们也一定生活在相近的时间(扩展数据图)。3 b).每组同时期个体与其他6个Chagyrskaya个体之间的遗传差异没有显著差异(Wilcoxon秩和检验,两者都是)P> 0.26)(补充表7.4).此外,这对同时期的父女在所有mtDNA序列中携带的差异数量最多,这意味着mtDNA多样性中不存在实质性的时间结构。综合起来,这些数据支持了这一假设,即所有11个Chagyrskaya尼安德特人都属于同一个群落。
这两具奥克拉德尼科夫遗骸彼此无关(p0= 1.14;95%置信区间,0.90-1.38),也与Chagyrskaya洞穴的任何个体无关。事实上,Chagyrskaya个体之间的成对遗传差异较低(p0= 1.0;95%置信区间,0.99-1.02)高于Chagyrskaya和Okladnikov洞穴个体间的差异(p0= 1.06;Wilcoxon rank-sum测试,P= 8.6 × 10−5(扩展数据图3和补充表7.3).这表明,奥克拉德尼科夫尼安德特人不属于查耶斯卡亚尼安德特人群落,该群落由我们获得DNA的11个个体所代表。然而,Okladnikov B的mtDNA与Chagyrskaya G的mtDNA是相同的。1 c).由于突变会随着时间积累,个体之间相同的mtDNA意味着这两个个体生活在彼此几千年的时间间隔内(补充表)3.9).此外,在此前发表的Chagyrskaya洞穴沉积物mtDNA样本中,38个样本中有2个更类似于Okladnikov A,而不是任何Chagyrskaya尼安德特人30..这表明,居住在两个洞穴的社区之间存在某种联系。
与其他人群的关系
为了探索Chagyrskaya和Okladnikov个体与其他尼安德特人的关系,我们调查了他们与之前发表的高质量尼安德特人基因组共享核苷酸变异的程度。所有13个新测序的个体都具有来自Chagyrskaya Cave (Chagyrskaya 8)的高覆盖基因组的大部分变异。5与Vindija洞穴(Vindija 33.19)约5万年前的尼安德特人基因组更相似3.与丹尼索瓦洞穴中9.1万至13万年前的阿尔泰尼安德特人(丹尼索瓦5号)相比2(扩展数据图。4).因此,尽管来自Chagyrskaya和Okladnikov洞穴的群落在基因上是不同的,但它们似乎都与欧洲尼安德特人有同样的关系,并且是同一尼安德特人群体的一部分;没有人显示出来自其他尼安德特人群体的近期基因流动的证据。
我们在这7个男性个体,3个尼安德特人,2个丹尼索万人和4个现代人类的Y染色体的6.9 Mb序列中确定了5416个变体(补充表)4.7).对于三个个体,我们只获得了低覆盖率序列(0.03- 0.3倍),而其他四个个体获得了更高的覆盖率(1.75- 42.2倍)(补充表)4.3).
我们构建了一个系统发育树,其中包括来自Chagyrskaya洞穴的4个高覆盖率的y染色体序列,以及其他3个尼安德特人、2个丹尼索万人和4个现代人的y染色体序列。1 d和补充表4.7).在尼安德特人中,所有四个Chagyrskaya序列都形成了一个分支,但它们更类似于El Sidrón 1253(西班牙),而不是地理上更接近的Mezmaiskaya 2(俄罗斯高加索北部)(图1)。1 d).这种地理结构的缺失与大约10万- 11.5万年前尼安德特人相当快速的扩张是一致的30..晚期欧洲的尼安德特人以及Chagyrskaya和Okladnikov尼安德特人都是这个种群的后代。
从剩下的三个个体中恢复的Y染色体序列数量不足以构建一个系统发育树,但在尼安德特人Y染色体彼此不同的位置,所有三个序列与其他查吉斯卡亚人Y染色体的派生变异多于与其他尼安德特人Y染色体的派生变异(补充表)4.9).
基于10个kb的窗口的覆盖率差异,我们检测到3个删除和5个重复(大小分别为20 - 2000 kb和10 - 200 kb)(补充表)4.4)在尼安德特人Y染色体上。最大的缺失是在Mezmaiskaya 2中发现的,它跨越了amely编码基因。因为蛋白质组学方法使用AMELY多肽的存在来确定骨头是否来自男性个体31,携带该基因缺失的雄性将被错误地分类为雌性(扩展数据图)。5).
mtDNA和Y染色体只跟踪单个位点,因此有必要通过常染色体遗传分析来研究基因流动的细节。在阿尔泰山区的尼安德特人和丹尼索瓦人之间的基因流动已经被观察到了,这个人(丹尼索瓦11)生活在7.9万- 11.8万年前,母亲是尼安德特人,父亲是丹尼索瓦人32.据估计,Chagyrskaya 8号中丹尼索瓦人血统的数量约为0.09%,混合事件发生在Chagyrskaya 8号生活的24,300±14,100年前33.为了研究混合的时间是否在其他Chagyrskaya个体中一致,我们寻找了他们基因组中与丹尼索瓦人基因组更相似的部分,而不是阿尔泰人或Vindija尼安德特人33.通过这种分析,我们在5个Chagyrskaya个体中确定了11个丹尼索瓦人祖先片段,这些个体的体长都超过0.2厘米(补充表)6.2).这些片段跨越3.2厘米(2.7 Mb),在Chagyrskaya A中发现的最长的1.5厘米(746 kb)。6.2).根据这些片段的长度,我们估计混合事件发生在Chagyrskaya个体生活的30000±18000年前,这与之前的估计是一致的(补充图。6.3).
丹尼索瓦洞穴被尼安德特人和丹尼索瓦人同时占领,大约在尼安德特人居住Chagyrskaya洞穴的同一时期34,35.然而,丹尼索瓦洞穴的石制品工业缺乏在查吉斯卡娅洞穴发现的Sibiryachikha变体的特征10.因此,尽管这两个洞穴距离很近,而且尼安德特人的母亲和丹尼索瓦人的父亲在丹尼索瓦洞穴中有一个后代,但在查耶尔斯卡娅洞穴被占领的几万年前就已经存在了25,我们没有发现任何证据表明,在Chagyrskaya个体存在之前的20,000年里,基因从丹尼索瓦人流向了Chagyrskaya尼安德特人(补充信息部分)6).
推断社会组织
我们利用8个个体(基因组覆盖超过0.9倍的个体)的纯合性基因组片段,研究了Chagyrskaya穴居人的群落和种群规模(补充信息部分)9).个体中较长的纯合性片段(大于10cm)意味着他们的父母在大约十代人之前拥有一个非常近的共同祖先,因此,很可能是一个小社区的一部分5,36.此外,具有中等长度纯合性片段(2.5-10 cM)的基因组的总体比例可以提供稍长的时间框架(约10-40代)内种群规模的信息。
先前对来自阿尔泰山脉的高覆盖率尼安德特人基因组的分析显示,约16.7%的丹尼索瓦5号基因组(参考文献。2)和Chagyrskaya 8基因组的19.3%(参考文献。5)具有中、长段纯合性。对这些模式的一种解释是,他们的父母是二级亲属2在不相关个体的背景下,在这种情况下,我们预计大多数其他个体有更少的纯合子片段。或者,这些数据可能是由于小的当地社区5,在这种情况下,所有的个体,除了最近的移民和他们的后代,将有广泛的纯合性片段。
在所有8个有足够覆盖的个体中,我们观察到1.6-14.9%的基因组具有长段纯合性,9.5-20.5%的基因组具有中间段纯合性(图5)。2和补充表9.2).我们注意到,这两个比例可能被低估了,因为在较低覆盖率下难以识别纯合度(补充表)9.1).因为我们在所有个体中都发现了大量的纯合子,所以我们得出结论,Chagyrskaya尼安德特人的当地社区规模很小。纯合子的数量也类似于在现代山地大猩猩基因组中发现的数量37(无花果。2 b),是一种濒危物种,生活在4-20只的小群落中38在该研究中,人们观察到二度亲缘关系的个体之间的交配是罕见的39.
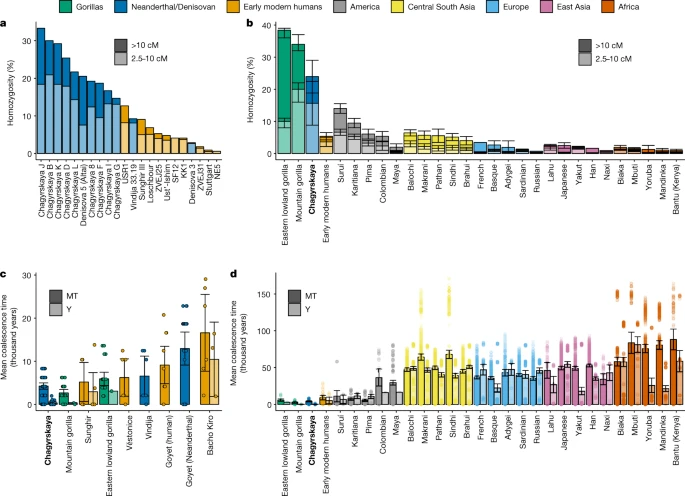
尼安德特人(蓝色)、早期现代人类(橙色)和现代大猩猩(绿色)在整个图中都是相同的颜色。现在的人口根据地理区域被着色(参见颜色键)。一个在古代个体(早期现代人、尼安德特人和丹尼索瓦人)中,长10厘米(深色)和2.5 - 10厘米(浅色)纯合子束中的基因组比例。b, Chagyrskaya穴居人,早期现代人(组合在一起)和现代人类和大猩猩种群的纯合子基因组的平均比例37.数据为平均值±95%置信区间的平均值估计。样本大小等于下面列出的mtDNA序列。c,尼安德特人、早期现代人类和大猩猩群体mtDNA (MT)和Y染色体的平均合并时间(分别为每对染色体的左右条)。d,早期现代人类(组合在一起)和现代人类和大猩猩种群的平均合并时间。c,d,数据为均值±95%置信区间,点均为两两比较。每个群体用于配对比较的Y染色体和mtdna -基因组数量如下:尼安德特人和丹尼索瓦人,Chagyrskaya (MT = 12, Y = 6), Vindija (MT = 4, Y = 0), Goyet(尼安德特人)(MT = 7, Y = 0);早期现代人类,Sunghir (MT = 4, Y = 4), vstonice (MT = 4, Y = 0), Goyet (MT = 5, Y = 0), Bacho Kiro (MT = 4, Y = 3),合计为(MT = 17, Y = 7);大猩猩、山地大猩猩(MT = 8, Y = 3)、东部低地大猩猩(MT = 7, Y = 2);美洲,Suruí (MT = 9, Y = 4), Karitiana (MT = 13, Y = 5), Pima (MT = 14, Y = 7),哥伦比亚(MT = 8, Y = 2),玛雅(MT = 22, Y = 2);南亚中部、俾路支(MT = 25, Y = 24)、马卡尼(MT = 26, Y = 20)、帕坦(MT = 25, Y = 19)、信德(MT = 25, Y = 20)、布拉希(MT = 26, Y = 25);欧洲、法国(MT = 29, Y = 11)、巴斯克(MT = 24, Y = 15)、阿代基(MT = 17, Y = 7)、撒丁岛(MT = 29, Y = 15)、俄罗斯(MT = 26, Y = 16);东亚、拉祜族(MT = 9, Y = 7)、日本人(MT = 28, Y = 19)、雅库特人(MT = 26, Y = 18)、汉族(MT = 34, Y = 15)、纳西族(MT = 9, Y = 6);非洲,比亚卡(MT = 23, Y = 22),姆布提(MT = 14, Y = 10),约鲁巴(MT = 23, Y = 11),曼丁卡(MT = 23, Y = 14),班图(肯尼亚)(MT = 12, Y = 10)。
为了进一步研究Chagyrskaya穴居人的社会组织,我们对比了11个母系遗传的mtDNA序列与6个父系遗传的y染色体序列的多样性。在无性别偏向过程的随机交配群体中,两个单亲本标记的平均结合时间是相同的。然而,观察到Y染色体的平均聚结时间(446年;95%置信区间113 - 1116年)显著低于线粒体基因组(4348年;95%置信区间,2043 - 6196年;Wilcoxon rank-sum测试,P= 4.1 × 105).在与47个现代人类和10个类人猿亚种的比较中,Chagyrskaya穴居人的y染色体与mtdna结合时间的比例是最低的,只有山地大猩猩的比例更极端(扩展数据图)。6).我们警告说,猿和尼安德特人的比例相似并不一定意味着这两个群体有相同的社会组织,因为这里有多个警告。首先,类人猿的数据非常多样化——例如,尽管有些类人猿出生在野外,但其他类人猿出生在圈养环境中(即人工群落),而且通常样本量非常小(补充表)8.1).第二,几种不同的情况可能导致相似的y染色体与mtdna的比例。这些因素包括:男性和女性世代时间的差异,男性后代分布的偏差(即,大部分孩子都是由一小部分男性生育的),以及女性偏好的迁徙。为了测试这些过程的相对重要性,我们模拟了这些场景的大量组合,拟合Y染色体和mtDNA的多样性及其与观测数据的比例(补充信息部分)8).我们使用模拟作为在观测数据95%置信区间内的模拟数据集的比例,对每个场景的可能性进行了近似计算。然后,我们使用赤aike信息标准(AIC)对不同的场景进行排名(补充表)8.5).
最佳拟合情景(AIC = 6.2)假设社区规模为20人,60-100%的女性是来自另一个社区的移民(补充表)8.4).然而,Chagyrskaya C和Chagyrskaya D之间共享的异质性表明,至少有一些雌性留在了它们出生时的群体中。如果只包含倾斜的后代分布,那么就不能很好地解释数据(AIC = 7.4),并且需要300个个体的大型社区规模。同时存在倾斜后代分布和雌性迁移的情况并不能提高仅假设迁移偏倚得到的拟合(AIC = 8.5)。仅包括世代时间差异的场景不适合数据(AIC = 8.5),并且需要看起来不现实的参数设置(例如,女性需要平均年龄为男性的两倍,补充表)8.4).此前对尼安德特人社区规模的估计从3到60人不等5,16,17,19在此范围内,最佳拟合情景包括女性迁移(补充图。8.4).这一结果表明,女性偏见的移民是Chagyrskaya尼安德特人社区社会组织的一个主要因素。
结论
我们展示了13个尼安德特人的基因数据,这是对尼安德特人群体进行的最大的基因研究之一。据我们所知,我们第一次记录了尼安德特人之间的家庭关系,包括一对父女。
所有个体的高度纯合性与山地大猩猩相似40这与生活在阿尔泰的尼安德特人的小社区一致。此外,根据Chagyrskaya和Okladnikov个体之间Y染色体的平均聚合时间比mtDNA短,以及共享mtDNA变体,我们认为这些小型尼安德特人社区主要由女性迁移联系在一起。
我们的研究结果提出了一个问题:阿尔泰社区的特征是否与他们位于已知尼安德特人活动范围最东端的孤立地理位置有关(特别是因为Vindija Cave的人口规模可能更大5),或者他们是否是更广泛的尼安德特人社区的特征。
因此,未来的研究应该在可能的情况下,从欧亚大陆其他地区的尼安德特人社区中取样,以进一步阐明我们最亲密的进化亲戚的社会组织。
数据可用性
每个图书馆的原始数据可在欧洲核苷酸档案登录号下获得PRJEB55327.所有样本和个体的BAM映射文件,VCF文件,一致的FASTA mtDNA序列和所有mtDNA的多重对齐都可以从这里下载http://ftp.eva.mpg.de/neandertal/ChagyrskayaOkladnikov/.
参考文献
格林,R. E.等。尼安德特人基因组序列草图。科学328, 710 - 722(2010)。
Prüfer, K.等。来自阿尔泰山脉的尼安德特人的完整基因组序列。自然50543-49(2014)。
Prüfer, K.等。克罗地亚Vindija洞穴的高覆盖率尼安德特人基因组。科学358, 655 - 658(2017)。
Hajdinjak, M.等。重建晚期尼安德特人的基因历史。自然555, 652 - 656(2018)。
Mafessoni, F.等。Chagyrskaya Cave的高覆盖率尼安德特人基因组。美国国家科学院学报。美国117, 15132 - 15136(2020)。
Peyrégne, S.等。两个早期尼安德特人的核DNA揭示了欧洲8万年的基因连续性。科学。睡觉。5eaaw5873(2019)。
Bokelmann, L.等。直布罗陀尼安德特人的基因分析。美国国家科学院学报。美国116, 15610 - 15615(2019)。
迈耶,M.等。中更新世司马德洛斯胡索古人类的核DNA序列。自然531, 504 - 507(2016)。
Derevianko, A. P.等。阿尔泰中部旧石器时代遗址恰杰斯卡娅洞的多学科研究(俄罗斯科学院西伯利亚分院考古与民族志研究所,2018)。
Kolobova, K. A.等。考古证据表明尼安德特人两次分散到西伯利亚南部。美国国家科学院学报。美国117, 2879 - 2885(2020)。
德列维安科、马尔金。Mustye Gornogo Altaya[阿尔泰山区的穆斯特人](Nauka, 1992)。
阿苏加,J. L.等。尼安德特人的根源:来自司马·德·洛斯·休索斯的头颅和年代证据。科学344, 1358 - 1363(2014)。
Higham, T.等人。尼安德特人消失的时间和时空模式。自然512, 306 - 309(2014)。
迈耶,M.等。一个古老丹尼索瓦人的高覆盖率基因组序列。科学338, 222 - 226(2012)。
灵长目动物社会系统的进化。Int。j·阿特拉通。23, 707 - 740(2002)。
Duveau, J., Berillon, G., Verna, C, Laisné, G. & Cliquet, D. Le Rozel(法国诺曼底)古人类脚印揭示的尼安德特人社会群体的组成。美国国家科学院学报。美国116, 19409 - 19414(2019)。
Mayoral, E.等。追踪伊比利亚海岸更新世晚期的尼安德特人。科学。代表。114103(2021)。
Vallverdú, J.等。古人类群体遗址结构内的睡眠活动区。咕咕叫。Anthropol。51, 137 - 145(2010)。
Lalueza-Fox, C.等人。尼安德特人群体父系交配行为的遗传证据。美国国家科学院学报。美国108, 250 - 253(2011)。
尼安德特人父系地域性的不确定证据。美国国家科学院学报。美国108(2011)英镑,共有财产占有一席之地。
Kolobova, K.等。来自Chagyrskaya洞穴(阿尔泰)的尼安德特人对自然环境的开发。Quartar667-31(2019)。
德列维安科,Markin, S. V. & Shunkov, M. V.阿尔泰旧石器时代中期的Sibiryachikha相。Archaeol。Ethnol。Anthropol。Euras。41, 89 - 103(2013)。
哈列维奇等人。阿尔泰西北部的新考古遗址(阿尔泰边疆区的Krasnoschekovsky和Soloneshensky区)。西伯利亚及其周边地区的考古学、民族志、人类学问题26, 263 - 270(2020)。
去除污染物:对分离单一化合物用于AMS年代测定的价值的重申。古代93, 1072 - 1075(2019)。
Slon, V.等人。尼安德特人的母亲和丹尼索瓦人的父亲的后代的基因组。自然561, 113 - 116(2018)。
彼得,M.等。尼安德特人和丹尼索瓦人Y染色体的进化史。科学369, 1653 - 1656(2020)。
Monroy Kuhn, J. M., Jakobsson, M. & Günther, T.估算史前种群的遗传亲缘关系。《公共科学图书馆•综合》13e0195491(2018)。
伊万诺夫,P. L.等。俄罗斯大公乔治·罗曼诺夫的线粒体DNA序列异质性确定了沙皇尼古拉二世遗骸的真实性。Nat,麝猫。12, 417 - 420(1996)。
斯图尔特,J. B.和Chinnery, P. F.线粒体DNA异质性的动力学:对人类健康和疾病的影响。Nat,启麝猫。16, 530 - 542(2015)。
Vernot, B.等。利用洞穴沉积物中的细胞核和线粒体DNA挖掘尼安德特人的种群历史。科学372eabf1667(2021)。
斯图尔特,格拉赫,R. F.,高兰,R. L.,格隆,K. J.和蒙哥马利,J.从牙釉质多肽中测定人类遗骸的性别。美国国家科学院学报。美国114, 13649 - 13654(2017)。
Douka, K.等人。丹尼索瓦洞穴古人类化石的年龄估计和上古石器时代的开始。自然565, 640 - 644(2019)。
彼得,b。m。阿尔泰山区尼安德特人和丹尼索瓦人之间10万年的基因流动。预印在bioRxivhttps://doi.org/10.1101/2020.03.13.990523(2020)。
Zavala, e.i.等人。更新世沉积物DNA揭示了丹尼索瓦洞穴古人类和动物群的更替。自然595, 399 - 403(2021)。
Jacobs, Z.等。西伯利亚南部丹尼索瓦洞穴的古人类占领时间。自然565, 594 - 599(2019)。
塞巴洛斯,乔西,P. K,克拉克,D. W,拉姆齐,M.和威尔逊,J. F.纯合子的运行:种群历史和性状结构的窗口。Nat,启麝猫。19, 220 - 234(2018)。
薛宇,王晓燕,王晓燕,等。山地大猩猩的基因组揭示了长期数量下降和近亲繁殖的影响。科学348, 242 - 245(2015)。
大猩猩社会组织的变异:生活史和社会生态学的观点。另一个星球。Anthropol。27, 218 - 233(2018)。
警惕,l等人。雌性习惯性分散的灵长类物种的繁殖竞争和近亲繁殖回避。Behav。生态。Sociobiol。69, 1163 - 1172(2015)。
Langergraber, K. E.等。黑猩猩群落有多古老?在高度父系社会中,y染色体最近的共同祖先出现了。j .的嗡嗡声。另一个星球。691 - 7(2014)。
Skoglund, P.等人。从西伯利亚尼安德特人体内的现代污染中分离内源性古DNA。美国国家科学院学报。美国111, 2229 - 2234(2014)。
安德鲁,R.等。人类线粒体DNA剑桥参考序列的再分析与修正。Nat,麝猫。23147(1999)。
Briggs, A. W.等。五个尼安德特人mtDNA基因组的靶向检索与分析。科学325, 318 - 321(2009)。
布朗,S.等。利用胶原蛋白指纹图谱和线粒体DNA分析鉴定西伯利亚丹尼索瓦洞穴的一种新的古人类骨骼。科学。代表。623559(2016)。
Douka, K.等人。丹尼索瓦洞穴古人类化石的年龄估计和上古石器时代的开始。自然565, 640 - 644(2019)。
索耶,S.等。两个丹尼索瓦人的细胞核和线粒体DNA序列。美国国家科学院学报。美国。112, 15696 - 15700(2015)。
克劳斯,J.等人。来自南西伯利亚的一种未知古人类的完整线粒体DNA基因组。自然464, 894 - 897(2010)。
Slon, V.等人。第四个丹尼索瓦人。科学。睡觉。3.e1700186(2017)。
迈耶,M.等。来自司马德洛斯胡索的一种古人类的线粒体基因组序列。自然505, 403 - 406(2014)。
Skaletsky, H.等人。人类Y染色体的男性特异性区域是一个离散序列类的马赛克。自然423, 825 - 837(2003)。
马利克,S.等。西蒙斯基因组多样性项目:来自142个不同人群的300个基因组。自然538, 201 - 206(2016)。
卡门,M.等。最近Y染色体多样性的瓶颈与全球文化的变化不约不同。基因组Res。25, 459 - 466(2015)。
门德斯,F. L.等。一个非裔美国人的父系谱系为人类Y染色体系统发育树增加了一个极其古老的根。点。j .的嗡嗡声。麝猫。92454 - 459 (2013);勘误表92637(2013)。
中提琴,B。中亚和西伯利亚的新古人类遗迹:最东端的尼安德特人?博士论文,维也纳大学(2009)。
克劳斯,J.等人。中亚和西伯利亚的尼安德特人。自然449, 902 - 904(2007)。
Mednikova, m . B。Postkranialnaya Morfologiya i Taksonomiya Predstavitelei Roda Homo iz Peschery Okladnikova na Altae(IAE SO RAN, Izd, 2011)。
确认
我们感谢D.卢卡斯和C.洛根对手稿早期版本的评论;H.用于计算机断层扫描的Temming;S. Sawyer和J. Krause钻出了Chagyrskaya 2号和Chagyrskaya 6号的骨头;R. Schultz帮助恢复样品的数字照片并上传主要数据文件。V.S.由阿隆奖学金资助,M.H.由居里夫人行动(MSCA-IF-EF-ST LIF)资助。欧洲研究理事会Horizon 2020研究与创新项目“RESOLUTION”(no. 844014)资助。欧洲研究理事会Horizon 2020研究与创新计划资助“FINDER”(编号:803147)。715069), R.G.R.获澳大利亚研究理事会奖学金FL130100116资助,T.H.获欧洲研究理事会第七框架计划(FP7/2007-2013)拨款324139“古时”资助,K.A.K.获俄罗斯科学基金会资助,项目N 21-18-00376, M.T.K.获波兰国家科学中心资助,项目2018/29/B/ST10/00906, B.V.获加拿大社会科学与人文研究理事会资助,Insight拨款435-2018-0943。该项目由欧洲研究理事会资助。694707到S. Pääbo)。
资金
马克斯·普朗克学会提供的开放获取基金。
作者信息
作者和联系
贡献
l.s.、S. Pääbo和B.M.P.设计了这项研究。A.P.D, S.V.M A.I.K.和a.k. K.A.K.收集样本。T.D。vs, M.H B.N, S.N。j。r,电子工程,M.G,其子as, P.K,特区,酸处理,T.H.和容积进行了实验室实验和/或分析。l . s . Peyregne,一般,L.N.M.I, T.D。vs, E.I.Z, M.H,美联社,S.G, A.B.M, D.H.L,特区,A.P.D, S.V.M,酸处理,a.k. K.D, M.T.K, R.G.R, T.H。B.V。A.I.K K.A.K.和B.M.P.进行分析。l . s . Peyregne,一般,L.N.M.I, T.D。vs, E.I.Z, M.H,美联社,A.B.M, D.H.L,酸处理,K.D, M.T.K, R.G.R, T.H。B.V。A.I.K, K.A.K, j.k.,答,Paabo和B.M.P.手稿输入从所有的作者写道。A.P.D, a.k., s.v.m., K.D, m.t.k., r.g.r., t.h., b.v., A.I.K.和K.A.K.提供了考古、地层和地质年代背景和解释。
相应的作者
道德声明
相互竞争的利益
作者声明没有竞争利益。
同行评审
同行审查的信息
自然感谢Krishna Veeramah, Katharine MacDonald和其他匿名审稿人对这项工作的同行评议做出的贡献。同行审查报告是可用的。
额外的信息
出版商的注意施普林格自然对出版的地图和机构附属的管辖权要求保持中立。
扩展的数据图和表
图1 Chagyrskaya和Okladnikov洞穴。
一个,南西伯利亚阿尔泰地区Chagyrskaya和Okladnikov洞穴位置图。的观点B,朝北的Chagyrskaya洞穴入口和C,朝南的Okladnikov洞穴入口。
图2 Chagyrskaya洞穴平面图及尼安德特人遗址位置。
一个,尼安德特人遗骸的空间分布。挖掘面积为灰色,蓝线(样带A-B)为中地层剖面的位置B.彩色的正方形和椭圆分别表示位于精确坐标或限定区域内的尼安德特人遗迹,并标注了相应的化石编号。B,沿A-B断面的地层剖面一个.尼安德特人遗骨的位置被投影到这个剖面上,所以每个化石不一定显示在它被复原的地层单元中。
图3 Chagyrskaya和Okladnikov之间的归一化两两差异。
一个,点表示两个剩余物之间的平均两两差异(y轴)(由所有剩余物对之间的中位数差异归一化)。被鉴定为相同的,一级和二级亲属的遗骸被命名(x轴表示第一个化石,数字表示第二个遗骸)。误差条是平均两两差异的100个自举估计的95%置信区间。水平线表示相同个体、一级关系、二级关系和不相关个体的预期归一化差异27.B,每个圆/正方形代表一个个体(蓝色代表Chagyrskaya,橙色代表Okladnikov),白色的小圆表示哪些遗骸来自这个个体。黑色的圆圈表示Chagyrskaya 8说明这块骨头的基因组序列之前发表过。正方形表示这个人是男性,圆圈表示这个人是女性。标记为一级亲缘、二级亲缘或共享异质系的个体。
图4古基因组间的变异共享。
错误条的中心显示表单的d统计值D((丹尼索瓦5/Vindija33.19),Chagyrskaya 8;Test,黑猩猩)和误差条是对应的95%置信区间,使用加权块折刀和5 Mb的块大小计算643,472个snp。| z得分| > 2的点用星号标注。虚线的垂直线在D = 0处。请注意,Chagyrskaya F是同一个人吗Chagyrskaya 8而且VindijaG1是同一个人吗出土的33.19.
扩展数据图5 .删除ameey染色体上的基因。
y染色体上1.8 Mb序列的缺失兹2(底部面板,浅灰色)相比Chagyrskaya D(顶部面板,不删除)。横轴表示在y染色体上的基因组位置,纵轴表示以10 kb为单位的覆盖率,由全染色体平均覆盖率归一化。Bin颜色表示人类参考y染色体上的区域类别,深色区域表示y染色体捕获阵列的覆盖范围。黑条表示已知的编码基因。
图6线粒体DNA与y染色体多样性的比值。
黑圈表示每个总体的平均估计,误差条是使用100次自举迭代对应的95%置信区间。负值表示y染色体多样性低于线粒体(mt) DNA多样性。
权利和权限
开放获取本文遵循创作共用署名4.0国际许可协议(Creative Commons Attribution 4.0 International License),该协议允许在任何媒体或格式中使用、分享、改编、分发和复制,只要您给予原作者和来源适当的署名,提供创作共用许可协议的链接,并说明是否有更改。本文中的图片或其他第三方材料包含在文章的创作共用许可中,除非在材料的信用额度中另有说明。如果材料不包含在文章的创作共用许可中,并且您的预期用途不被法律法规允许或超出了允许的使用范围,您将需要直接从版权所有者那里获得许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/by/4.0/.
关于这篇文章

引用这篇文章
斯科夫,Peyrégne, S。et al。对尼安德特人社会组织的基因洞察。自然610, 519 - 525(2022)。https://doi.org/10.1038/s41586-022-05283-y
收到了:
接受:
发表:
发行日期:
DOI:https://doi.org/10.1038/s41586-022-05283-y
这篇文章被引用
在西伯利亚洞穴中发现的第一个尼安德特人家族
自然(2022)
