








文摘
鱼尾弹点是最早广泛抛物型在南美,并分享年表和techno-morphology克洛维,北美最古老的弹体类型。两人都暂时与晚更新世megafaunal灭绝。尽管人类利用巨型动物的难以捉摸的直接证据在南美洲一直鱼尾灭绝的辩论中,最近的一篇论文显示强大的巨型动物的时间密度和空间分布之间的关系和鱼尾弹点,并提出这种武器设计和用于巨型动物狩猎,导致它们的灭绝。如果是这样的话,这项技术必须从post-FPP技术(即截然不同。,early Holocene projectile points), used for hunting smaller prey, in terms of distribution and functional properties. In this paper, we explore the changes in projectile point technology, as well as the body mass of potential megafaunal prey, and show that Fishtails were strongly related to the largest extinct megafaunal species.
介绍
鱼尾弹点(FPP)代表最早广泛岩屑类型在南美洲的工具1,2,3,4,5。这些点最初被定义为“下降点”朱尼厄斯鸟恢复一组他们在更新世晚期文化层次的下降的洞穴,在巴塔哥尼亚的南端6。这个考古复杂成为众所周知的主要因为它是几世纪后在年龄和与北美克洛维斯点共享一些techno-morphological特性1,7,最早广泛techno-complex整个美洲8,9,10。鱼尾传统意义上被定义为一个没有点宽三角形或披针形叶片,凸边,宽厚的肩膀6,7。虽然近几十年来的研究表明重大FPP之间的形态变异,大部分标本分享特定的设计特性,比如广泛而薄刀片,并且通常有凹槽的茎7。鱼尾和克洛维斯点也常与megafaunal仍然存在。时间和相似风格的克洛维斯和鱼尾弹点了一些研究者所代表的合理建议的表达同样的文化或技术的现象1,7,11,12,也都与megafaunal打猎。尽管不同意见,克洛维斯,在较小程度上,FPP一直争论的核心早期美洲的。
在北美,克洛维斯点之间的联系和巨型动物的狩猎是相当清楚的13和造就了我们长期和持续的辩论在更新世megafaunal灭绝人类的角色14,15,16,17,18。尽管这场争论尚未发生在南美洲由于有限的人类利用巨型动物的直接证据19,20.,21,最近的一篇论文给FPP更坚定和更重要的角色在灭绝的争论22。基于间接证据,时间和空间之间的相关性FPP和巨型动物,睡午觉,佩雷斯22建议这些弹点是用于狩猎megafaunal物种的主要武器。特别,他们表明,(1)FPP同时出现在南美洲南部,ca。13 k年卡尔英国石油(BP)和类似的地形,开放地区巨型动物的密度也达到最高的价值观,和(2)FPP和巨型动物几乎完全消失11 k年后卡尔BP。这些结果表明,FPP的迅速和成功的传播技术推动了高人口增长率在晚更新世23然后,contributed-along与其他环境因素和间接影响24——巨型动物的灭绝。
使用FPP亨特巨型动物可以通过分层评估证据水平:(1)直接证据(协会杀死/屠杀地点),(2)功能的证据(独特的特征,使得他们专门为狩猎巨型动物),和(3)相关证据(空间和时间相关性和巨型动物)。假设找到直接证据并不是一个必要的巨型动物狩猎的期望25,26和相关证据已被评估22在本文中,我们评估FPP捕猎巨型动物的功能特征。如果人类是主要的代理负责灭绝巨型动物的影响显著,后突然出现的FPP (ca。13 k年BP)22,这种技术必须被视为一个真正的革命行为27。因此,我们期望强大功能之间的关系FPP和megafaunal打猎。如果是,FPP必须明显不同于其他早期弹点不用于巨型动物狩猎、分布和功能性质。在本文中,我们解决这些问题,以便更好地理解之间的关系的改变弹点技术和潜在megafaunal猎物(体重44公斤)14人类的猎人。首先,我们估计和比较早期的时间和空间分布弹点可靠日期最新的更新世(ca。13 k-11 k年卡尔BP)和全新世早期(11 k - 8.5 k年卡尔BP)在南美洲的南锥(即。,FPP Tigre支付帕索,推拿,Ayampitin和巴塔哥尼亚三角形)FPP空间集中的地方。为此,我们使用站点分布和放射性碳年代测定与最大熵分布模型(MaxEnt)28,29日和总结概率分布(SCPD是)方法30.,31日。如果FPP专门为狩猎巨型动物灭绝22从时间和地理的角度来看,我们期望他们所取代,灭绝和所有对其色散区域后,其他类型的分低效率的狩猎大型哺乳动物(例如,越来越少的杀伤力)。然后我们探索不同类型的早期弹点的设计特点及其关系megafaunal打猎。为此,我们分析形态学和早期弹点的技术数据类型的南锥。特别是,我们评估弹丸的morpho-functional对应点与可用的潜在megafaunal猎物不同时期(最新更新世和全新世早期)和地区(草原、安第斯山脉和巴塔哥尼亚)。我们测量设计有效性(健壮性和渗透),破坏能力,杀伤力,每个点类型的制造成本32,33,34,35,36,37。如果FPP是人类成功的关键技术因素亨特巨型动物开始在13 k年卡尔BP,我们预计,从morpho-functional的角度来看,这些点来显示组织损伤的最高能力。我们的结果加强预期的变化在南美洲南部早期弹点技术是与体重的变化可用的猎物。FPP灭绝巨型动物捕食,密切相关,与全新世早期点,他们的设计被要求限制捕食大型动物。
结果
时间密度的空间分布和变化弹点类型
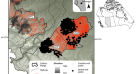
弹点的分布的最新更新世和全新世早期从南美洲南部画使用空间分布建模MaxEnt方法实现28,29日。我们认为弹点ca的记录。骁将k卡尔BP(最新更新世)和11 - 8.5 k卡尔BP(全新世早期),以及生物气候变量PaleoClim中可用38为每一个时期。图1显示了不同类型的抛射体的空间分布分两个时间。MaxEnt模型的预测能力高的所有类型,AUC值在0.8和0.99之间(FPP-Andes: 0.800;FPP-Pampas: 0.897;FPP-Patagonia: 0.973;推拿治疗:0.978;Ayampitin: 0.797;巴塔哥尼亚三角形:0.989),这表明我们获得很好的优秀的估计每个弹点的分布类型。分布地图显示,最新的更新世FPP显示高值的电位分布和大量的记录在潘帕斯草原,有分散出现在巴塔哥尼亚;Tigre点限制在潘帕斯草原的几个地点。在全新世早期,我们观察到巴塔哥尼亚三角形弹点FPP-Patagonia类型呈现相似的分布; Tuina and Ayampitin are superimposed on FPP-Andes, and Pay Paso occupy almost the same sites as Tigre points (Fig.1)。

南美南部弹点的空间分布。地图显示最新的更新世(电位分布的一个)和全新世早期弹点(b)。颜色代表潜力分布值高于0.6(浅色)和0.8(强烈的颜色)。Tigre和支付帕索弹点只有网站画,因为小样本大小。南美北部的网站鱼尾弹点也画黑色小点。地图生成和QGIS 3.16“汉诺威”(https://qgis.org/en/site/index.html)。
时间的变化在不同类型的弹点的密度进行评估使用放射性碳年代测定和总结概率分布方法(SCPD是方法30.,31日)。图2表明,密度的变化日期FPP几乎同步的安第斯山脉,草原,和巴塔哥尼亚,显示一个明确的增加经过13 k卡尔BP和之间的强烈下降12.5 k和11 k卡尔BP。Tigre点与去年同期记录FPP的潘帕斯草原(无花果。2)。全新世早期弹类型出现在11 k卡尔BP潘帕斯草原(支付帕索)和巴塔哥尼亚(巴塔哥尼亚三角形),后者地区与后来的日期。在安第斯山脉,推拿分部分重叠FPP(无花果。2)。然而,这种叠加是由于一些老日期从一个网站(令人惊叹印加平图拉斯河手洞4),像离群值。

总结概率分布曲线在南美洲南部三个区域。安第斯山脉的时间变化显示(一个)、南美大草原(b)和巴塔哥尼亚(c),使用每个弹点的估计SCPD是曲线类型或风格。我们只包括弹点类型日期为8.5 k以上年卡尔BP。
弹点设计,功能组织损伤和杀伤力
我们探讨了不同弹点设计在时间和地理空间使用宽度和厚度32,横截面周长(TCSP视为代表组织损伤)33,34伤口面积(WSA)36,投资在制造业(WI)37。图3显示厚度和宽度之间的关系在对数刻度平均值的六个研究抛物点类型,分别考虑FPP从安第斯山脉,草原和巴塔哥尼亚。这些变量之间的关系是由最小二乘回归直线。通过增加弹点的宽度,潜在的猎物增加造成的损害;和减少厚度,渗透能力增加,也打破由于稳健性损失的风险。因此,有必要补偿宽度和厚度,使弹丸穿透点维护能力,导致严重的伤害和保持的32。由于最小二乘回归估计在对数刻度线,这两个变量之间的关系是指数(图。3)。

散点图的厚度和宽度之间的关系在对数刻度分别六弹点类型。实线是最小二乘回归的数据点代表平均值为每一个弹点类型。灰色区域表示置信区间的最小平方线。弹点意味着在回归线上的左上角显示设计,最大化的鲁棒性,并指出在回归线下面的右下方显示设计,最大化渗透。的横截面周长(TCSP)和工作投资(WI)也显示在颜色梯度(值用毫米表示)和球的大小,分别。看到布坎南和汉密尔顿32为类似的方法探索有效性和破坏能力。
图3还显示最新的安第斯山脉的更新世弹点和潘帕斯草原最大的宽度值,而全新世早期点显示最低的值。潘帕斯草原的FPP和安第斯山脉被放置在最小二乘直线附近,表明他们为了最大化的鲁棒性和穿透属性32。FPP的潘帕斯草原和安第斯山脉和Tigre弹点显示更高价值的TCSP反映更大的能力来生成组织损伤(无花果。3)。全新世早期弹点来自潘帕斯草原和安第斯山脉呈现显著降低值TCSP比最新的更新世(表1),建议减少生成组织损伤的能力随着时间的推移,即使这是狭隘的平衡通过增加渗透。这种差异不是在巴塔哥尼亚(表1),鱼尾和全新世早期点(巴塔哥尼亚三角形)显示类似的价值观TCSP(无花果。3)。morphospace的无花果。3,全新世早期设计显示更多random-like pattern-far从最小二乘回归比最新的更新世,表明没有一个因素限制了设计。
伤口面积(WSA),定义为TCSP和穿透深度的产物36措施的杀伤力,胸腔的一种动物。这对FPP指数计算,Tigre推拿,Ayampitin,支付帕索弹点。我们的结果表明,所有的晚更新世的设计,除了FPP从巴塔哥尼亚,可能更致命的比早期的全新世(表2)。
最后,投资估算工作(WI)(和修改取自Aschero Hocsman37)在无花果。3,我们赋值从1到4为不同的技术程序(如有两面的减少或变薄),根据工作难度增加。FPP WI(得分4)最高,其次是Tigre(得分3),巴塔哥尼亚三角形,Ayampitin并支付帕索(分2),推拿弹分(分数1)(图3)。Techno-morphological FPP的特点(伟大的宽度,厚度最小,有两面的变薄,打捞筒剥落和开槽)必须在塑造过程中要求工作的更大的投资。较小的工作是参与到全新世早期的设计,需要较小的叶片宽度和简单有两面的还原/变薄。
身体质量潜在的猎物
我们描述了体重变化通过时间和空间megafaunal物种与人类有关的考古上下文或严格使用以前的基于回归模型的估计40。图4表明体型值随地区。最新的更新世物种在安第斯山脉(Notiomastodon platensis, Hippidion devillei喇嘛guanicoe,喇嘛(骆马)股薄肌)是比全新世早期的重约1100公斤(喇嘛guanicoe,Hippocamelus antisensis,骆马骆马)。在潘帕斯草原,最新的更新世物种(大地懒属americanum (m),科仕neogeus Glossotherium robustum,喇嘛guanicoe, Mylodon darwini)约有950公斤重比全新世早期(喇嘛guanicoe和Ozotoceros bezoarticus前(),在巴塔哥尼亚Mylodon darwini,喇嘛guanicoe,喇嘛(骆马)股薄肌,Hippidion saldiasi)比后者重约250公斤(喇嘛guanicoe和Hippocamelus bisulcus)。这种模式的最新体重差异更新世和全新世早期动物类似于上述的差异弹点设计有效性(健壮性和渗透),破坏能力,和制造成本,在安第斯山脉上的巨大差异和潘帕斯草原,和小的巴塔哥尼亚(表1和无花果。4)。

体型变化megafaunal物种被人类。箱形图的巨型动物的体重(公斤)地区和时期。图显示的最小值和最大值(须),第一和第三个四分位数(边界框),和中位数(中线)。
讨论
弹点的多样性在时间和空间
尽管non-preserved磨木矛或骨性弹提示可能被用于狩猎41,42、鱼尾石点是最早广泛弹点型在南美洲(骁将k卡尔BP)和唯一分布在所有的纬度和大部分环境中,最低的密度在热带低地和最高的南锥(无花果。1和2)。更高的密度在这些地区由开放环境中观察到的大草原和热带稀树草原的更新世,尤其是草原和巴塔哥尼亚43,更大的多样性和丰富的巨型动物也被记录下来44。如果FPP专门为狩猎巨型动物灭绝22,我们预计他们会灭绝后代替了其他类型的弹点,更小的和更没有效率的狩猎大型哺乳动物,在他们的色散区域。如图所示,布坎南等。45更大的北美Paleoindian点被用来狩猎大型猎物和小点被用来狩猎小猎物。
FPP替换后的期望巨型动物灭绝是实现整个南锥,一般来说,和三个地区(南部的安第斯山脉、草原和巴塔哥尼亚),特别是(图。2)。在所有的三个区域,FPP出现约13 k年卡尔BP,之前megafaunal物种的数量和密度的增长突然停了下来,开始显著下降12.9 k和12.8 k年卡尔BP22。从12 k年卡尔BP开始,其他类型的弹点出现在安第斯山脉(推拿)和潘帕斯草原(Tigre并支付帕索),完全取代FPP 11 k年卡尔BP,当时巨型动物几乎消失了。在巴塔哥尼亚,虽然FPP消失早于其他地区(11.5 k前不久年卡尔BP),有大约500年的时间没有记录的标准化弹点。有趣的是,在这个地区,巨型动物灭绝之后,南美也出现回落46,47和减速下降,人口的增长似乎是更加剧烈,超过其他地区23。所以,抛射点在该地区的长期隐身也可能产生影响的人口密度低和/或减少可用性后的哺乳动物灭绝。只有大约11 k年卡尔BP中型三角形弹点出现在巴塔哥尼亚,然后在大部分地区存在几千年(无花果。2)48。
尽管FPP最早和最广泛的抛射点在南美洲南部,他们并不是唯一最新的更新世时代。有几种其他类型当代巨型动物,但较小的空间和时间色散比FPP(无花果。1和2)。首先,Tigre点,限制分散在乌拉圭的潘帕斯草原,显示部分时间重叠FPP和一起消失在ca。11 k年卡尔BP。在最近的一篇论文,提出了与FPP non-stemmed三角点是可能的潘帕斯草原49,但是我们没有分析他们在这里因为他们安全地尚未过时了。唯一可用的日期(ca。8 k年卡尔BP)来自阿罗约瑞士2(草原),但这些点与人类遗骸,主要是嵌入到骨骼50,因此,他们可能不会用于狩猎或者至少他们不是专门用于狩猎。相反,Tigre点暂时的增加伴随着FPP的衰落(12 k−11.1 k年卡尔BP)51(无花果。2)。所以它不能排除,他们代表着当地的变异点也为巨型动物狩猎,他们被废弃了,消失,FPP一样,在晚更新世灭绝。
不同的情况下是推拿弹点分布在南部的安第斯山脉。虽然在时间和FPP推拿分部分重叠(和巨型动物),这些点的更新世时代不应该被认为是可靠的,因为两个古老的日期(令人惊叹来自印加平图拉斯河手洞4)似乎离群值。推拿治疗点,不像Tigre明显增加的频率在全新世早期,大多数megafaunal物种已经灭绝,之后FPP消失(无花果。1和2)。这时间趋势与考古资料表明,推拿点通常是与现存的物种,特别是喇嘛骆马52。推拿弹点应该没有相关megafaunal狩猎,但仍可能被用于狩猎现存骆驼科,之前(如果几个例外日期是有效的),在更新世灭绝之后。
另一个最新的更新世抛物型,除了Tigre推拿治疗,但大部分位于研究区之外,El Jobo披针形点,比FPP的北部和安第斯山脉的分布。虽然表面上保存完好,El Jobo点很少,最古老的日期从Taima Taima网站(ca。16 k年卡尔BP53)是有争议的23;他们可能与FPP准同生。超过这个时间问题,El Jobo分和巨型动物之间的关系(乳齿象和大地懒属)54,55强,因此,他们可能是当地的一个类型的变体矛头用于杀死的猎物。然而,自El Jobo点只有轻微的地理联系Pampean megafaunal物种和重叠更与北方物种特别是长鼻目动物——比FPP,尚不清楚El Jobo和FPP同步地理变异的弹点类似的功能,或如果是前者被后者所取代。实验的茧衣56,57显示,克洛维斯点(类似于FPP)有效地渗透到象隐藏,和玛瑙盆点(更类似于El Jobo58)似乎更适合冲通过野牛或牛皮。如果是这样的话,效果可以解释更换El Jobo FPP。
Morpho-functional早期弹点的属性
我们的期望是,如果FPP是人类成功的关键技术因素亨特巨型动物开始在13 k年卡尔BP,这些点应该显示功能性能的最高水平。这是因为大型动物通常更难猎杀,和需要更有效的和致命的武器比用于小游戏45,59。抛射体的主要功能点是打开一个伤口在动物的皮肤和组织,渗透到猎物造成大量出血,并到达一些重要器官,造成快速死亡或严重伤害,让猎人去追求和完成受伤的动物32,45。
我们的研究结果显示,最古老的类型的碎石头弹点的南美,一般来说,和FPP,特别是,更大,更有害的,致命的,技术上更复杂的比大多数早期的全新设计。显示在无花果。5,潘帕斯草原的FPP和安第斯山脉,从乌拉圭Tigre点,他们暂时和空间相关的巨型动物,那些TCSP最高的价值,因此,最高的组织损伤的潜在能力。损害的大小主要与弹点的最大宽度,更快的提高会导致更大的冲击和出血的猎物59。虽然TCSP指数与渗透能力呈负相关,和减少弹点的大小会增加渗透,这是限制的需要为快速出血导致受伤的猎物60。因此,它是至关重要的实现之间的平衡实现充分渗透TCSP足够小,大到足以产生一个伤口,容易出血36,60。

箱形图的TCSP指数弹点类型(值表示在毫米),显示每种类型的典型形式的顶部轴。图显示的最小值和最大值(须),第一和第三个四分位数(边界框),和中位数(中线)。弹点的功能分配是基于人种学和实验数据60。
安第斯山脉和潘帕斯草原FPP还提供了一个更高效的平衡所代表的宽度和厚度(最小二乘在无花果。3)。点的厚度减少,鲁棒性降低和脆弱性增加,使它变得不那么耐机械力的影响,目前对猎物的影响,比如屈曲。因此,布坎南和汉密尔顿32说了Paleoindian弹点,似乎FPP被设计为长,宽,和薄,罢工之间的平衡的渗透力量最大化和风险最小化的矛头打破32。相似的影响。伤口面积分析(表2),清楚地表明,FPP技术是一种高效的设计具有高伤害能力。一般来说,点之前12000年卡尔BP将产生伤口比全新世早期的12%至24%。
通过比较最新的更新世弹技术在大陆范围内,超过我们的研究区域,三个额外的趋势出现。首先,El Jobo点,潜在的古老南美弹技术,和为数不多的类型与巨型动物,有较小的伤害能力比南部Tigre和FPP类型(图。5)。如果El Jobo的确是南美洲最古老的弹点类型,它们可以被用于狩猎巨型动物,直到被FPP取代,可能一个新的,更有效,更可靠的武器狩猎大型哺乳动物。其次,北美弹点更清楚地与巨型动物(例如,克洛维)也会有更大的潜力相比,组织损伤后北美弹类型(例如,福尔松的)(无花果。559)。这种模式类似于观察到在南美洲南部,但损害的价值能力的北美点不到Tigre和FPP的估计。它也是有趣的,看看点似乎变得越来越宽体,narrow-stemmed向南(见图。5)与直边克洛维斯点。
最后,大小和TCSP弹点的变化可能与使用不同的武器系统,从人种学的集合比较弹点TCSPs提出的武器系统在休斯责任实验武器(表四61年)。由图所示。5,大多数最新的更新世和全新世早期弹类型从北美和南美都可能使用梭标投射器飞镖61年通过实验研究,提出了62年。只有最大的类型的潘帕斯草原(Tigre但特别FPP)似乎兼容投掷或把长矛63年,64年。后者武器系统通常是与狩猎大型猎物65年并将适当的地区巨型动物更加丰富和多样化。虽然已经提出,Paleoindian弹点可能是刀子12,66年骨折,实验研究表明,鱼尾点主要是用作炮弹62年。此外,一种点的主要功能是一个弹丸并不意味着它不能也被用作一个刀67年。
除了最新的事实更新世弹点从南美洲南部提供最高水平的性能,他们的高档功能性属性也需要很高的专业知识和生产成本(无花果。3)。FPP flintknappers产生点的宽度和最小厚度,使用的技术包括有两面的变薄,打捞筒剥落,通常开槽,必须要求一个更大的工作和更多的技能相比,成本需要生产其他弹丸的设计7。点宽,薄刀片将导致更多的组织损伤的猎物,这可能抵消了更大的工作和技能投入生产。我们似乎是合理的,这种专门的武器已经取代了以前的和低效率的设计(例如,El Jobo),但将不再生产一旦成为不必要的昂贵物品的猎物后设计(巨型动物)消失。事实上,大多数早期的全新世弹点类型,除了少伤害能力,要求更低的制造成本和flintknapper技能比生产FPP的必要。特别是,他们破坏能力低于鱼尾和Tigre点(表2和无花果。3)由于其较小的叶片宽度,尤其是推拿和支付帕索类型。尽管大多数早期的全新类型的健壮性与晚更新世的,有利于渗透(图的最小厚度。3),他们的伤害能力低20%(表2)。显示在无花果。3工作Ayampitin投资,支付帕索,巴塔哥尼亚三角形,和推拿弹点仅限于有两面的还原/变薄,和较小的叶宽生产68年,69年,70年,71年。
我们期待更好的狩猎FPP的性能比其他设计通常是满足,除了Tigre点礼物,一起潘帕FPP,最高的伤害能力值。加上Tigre点共存的事实和巨型动物消失后他们灭绝,表明这些点也可以被用来狩猎megafaunal物种。Tigre点厚和有一个较小的渗透能力比FPP(无花果。3),因为它们提供一个低效率的宽度和厚度之间的关系。他们可能需要更少的制造成本和flintknapper的技能比FPP由于稀释的较小的工作和开槽的缺失。虽然Tigre点可能只有一个本地最新更新世的文体变异点,增加频率暂时恰逢FPP的衰落(12 k - 11.1 k年卡尔BP)70年(无花果。2)和megafaunal丰度和多样性的减少12 k年后卡尔BP。因此,另一种解释可能是,猎物的变化可用性后12 k卡尔BP的高性能FPP不必要和Tigre点更可行的(便宜)技术替代狩猎巨型动物和较小的猎物。
潜在的减少大小捕杀猎物在南美洲南部最新更新世至全新世早期反映在哺乳动物的平均体重减少考古遗址(无花果。4)。如果早期弹点类型多样与这种变化,不仅是组织损伤的能力点后将减少巨型动物灭绝,还这个减少的大小应该符合体重减少的大小在每个地区。如无花果所示。6以来我们的结果支持这一假设减少TCSP值下降值与中间区域动物的体型。TCSP值显著或显著的差异为安第斯山脉和潘帕斯草原,但巴塔哥尼亚(表1),比如从观察到的模式预计将在我们的结果。巴塔哥尼亚FPP表现出相对较低的伤害能力,也是一个低效宽度和厚度之间的调整,甚至对巴塔哥尼亚相比早全新世点(巴塔哥尼亚三角形)。即便如此,这种模式并不一定与预期相反,因为猎物的身体质量的变化在巴塔哥尼亚灭绝之后似乎已经不太明显,必然会导致只有小猎物之间的平均大小的差异。这可能是因为最新更新世巴塔哥尼亚巨型动物相对较小而Pampean和安第斯山脉,也因为早全新世潜在的猎物(喇嘛guanicoe和Hippocamelus bisulcus)相对比那些从安第斯山脉或潘帕斯草原。

箱形图的TCSP弹点类型和身体质量指数的巨型动物按地区和时期,在z分数数据规模。图显示的最小值和最大值(须),第一和第三个四分位数(边界框),和中位数(中线)。地图代表物种多样性和弹点分布生成QGIS 3.16“汉诺威”(https://qgis.org/en/site/index.html)。
总之,自20世纪中叶以来,鱼尾弹点已经密不可分的争论南美的早些时候工作;主要是因为他们的早期年表,形态与克洛维斯点,亲和力与巨型动物及其时间重叠。不像在北美,鲜有证据表明人类开发南美的巨型动物几乎已经无效的争论在人类和最新的更新世灭绝之间的关系。尽管最近的一项研究显示,一个强大的物种灭绝之间的相关性和鱼尾分和推断人类的崛起22鱼尾,功能相关性分和巨型动物狩猎到目前为止还没有评估。我们已经表明,首先,改变morpho-functional早期南美弹点的属性变化在时间和空间上与身体大小的捕杀猎物。其次,鱼尾点不仅暂时和空间与巨型动物灭绝,而且他们最有效的技术来捕食他们由于更大的组织损伤的能力,损伤/渗透比率和杀伤力。第三,鱼尾点的提高效率也使他们在技术上更昂贵(由于有两面的变薄,打捞筒剥落,通常开槽)。第四,鱼尾代表一个新的、革命性的技术,可能从北方引进7这是更好的狩猎和特别设计的巨型动物。第五,巨型动物的灭绝后,鱼尾点取代南美洲南部不同地区的其他类型的点,少破坏能力,损伤/渗透比率,和杀伤力,但技术更便宜,即使有效的杀死足够可用的小猎物。最后,尽管直接考古证据的人类开发南美的巨型动物仍然是难以捉摸的19,我们的研究结果揭示了核心作用FPP的巨型动物狩猎和加强假设人类有直接和显著的影响在其灭绝。未来的工作需要澄清重磨如何影响FPP的功能性质,FPP技术从何而来,并评估潜在的捕食策略和武器系统之间的联系和不同种类的狩猎的猎物(大小和行为学)。
材料和方法
不同类型的弹点的空间分布类型最新的更新世和全新世早期从南美洲南部使用最大熵估计建模方法实现MaxEnt方法28,29日。这种方法被广泛用于古生态学和考古学,因为它是健壮的和基于你们那里的数据28,29日,72年,73年。弹点的数据用于分析由记录绝对日期或高度可靠相对日期和精确的地理位置。这些数据是来自原始出版物和最近的一些编译(数据S1),尽管这可能反映了偏见的研究工作和/或系统性考古记录的可见性影响因素,样本的代表性不妥协。MaxEnt分析,我们单独考虑记录不同类型的弹点分配给最近的更新世(ca。13 k和11 k卡尔BP)和全新世早期(ca。11 k和8.5 k卡尔BP), FPP的地理变异,以及用作预测中可用的生物气候变量PaleoClim每个时期(http://www.paleoclim.org/)38。模型的性能估计为每个弹点类型使用AUC统计(即评估。曲线下的面积)28,29日衡量模型的适合不同在0和1之间(更好的预测价值)值低于0.50,表明该模型并不比那些获得的机会和价值高于0.80显示非常好或优秀的适合。电位分布的地图都是画在QGIS 3.16.4汉诺威的74年。
颞密度的变化弹点的不同类型的评估使用放射性碳年代测定和总结校准概率密度法(SCPD是方法)30.,31日。FPP的放射性碳年代测定使用的是编制和佩雷斯睡午觉22和其他的数据集是补充日期弹点从考古出版物(数据类型S2)。这种放射性碳数据集生成使用标准的验证标准提出了考古学、睡午觉这样总结等。23。放射性碳年代测定是校准使用南半球SHCal 20曲线。SCPD是曲线估计为每个弹点类型使用校准日期被网站200年的间隔年,然后使用移动平均平滑窗口大小的500年。放射性碳年代测定的校准和SCPD是曲线估计方案rcarbon4.0 R软件75年。因为我们是弹技术感兴趣发生在次megafaunal灭绝,我们限制分析弹点类型的最早记录日期是8.5 k以上年卡尔BP。
弹点设计和能力组织损伤使用形态学和技术数据(数据进行了探讨S3)。我们获得度量数据(宽度和厚度)127 FPP从南美洲南部和另一个303年早期的点。弹点包含在分析完整或展出骨折,并没有阻止他们测量的宽度和厚度。那些叶片再生影响的人排除在数据库(数据S3)。后,这是因为这些点的宽度的减少,导致大量出血的主要功能可以不再已正确实现,所以他们可能丢弃或作为刀,重新提出了FPP和不对称的叶片63年。我们描述弹点类型之间的差异和地理变异FPP使用宽度和厚度32提示的横截面周长(TCSP27,33,61年),工作投资制造业(WI)37。我们第一次的厚度和宽度之间的关系进行了探讨六弹点分别types-considering安第斯,Pampean,巴塔哥尼亚鱼尾抛物点对数刻度,使用最小二乘回归方法。执行这个分析布坎南和汉密尔顿32使用经典的异速生长的方法,探索弹点的鲁棒性和渗透的差异。其次,我们探索组织损伤的鉴别能力TCSP使用作为一个代理27,61年。TCSP来自弹道学,计算的最大宽度和厚度弹点叶片(TCSP = 4×√(宽/ 2)2+(厚度/ 2)2),可以被认为是一个代理,直接正比于其损害的能力27,33,61年。我们也计算了WSA指数(伤口面积= TCSP×穿透深度),目的是评估点的杀伤力36。穿透深度能力为每个类型的弹点是根据实验数据计算TCSP和穿透深度之间的关系,通过Sitton et al .(图。4)39。
TCSP变异和WSA每个弹点类型与体重估计megafaunal物种的差异40,76年与人类相关的来自不同地区的考古记录(安第斯山脉,南美大草原和巴塔哥尼亚)和时间(最新更新世和全新世早期)。最后,工作的估计投资(WI)为每个类型是基于Aschero和Hocsman弹点37。我们分配一个值从1到4为不同的技术程序(如有两面的减少或变薄),根据工作难度,增加订单。投资工作要求如下:如果让弹点包括有两面的减少(1分),有两面的变薄(分2),打捞筒剥落(得分3),和开槽(得分4)。
数据可用性
纸张内的所有相关数据和辅助数据文件。
引用
明天,j . e . &明天,t . A .地理变化槽弹点:一个半球的视角。点。Antiq。64年(2),215 - 230 (1999)。
politi g·克洛维斯。在起源和适应性(eds Bonnichsen r & Turnmire k) 287 - 301(德州农工大学出版社,1991年)。
水域,m R。,Amorosi, T. & Stafford, T. W. Redating Fell’s cave, Chile and the chronological placement of the Fishtail projectile point.点。Antiq。80年,376 - 386 (2015)。
Hermo D。,Miotti, L。&Terranova, E. Exploring Technological choices in fishtail points from southern contexts: A comparative overview.PaleoAmerica8(1),79 - 94 (2022)。
鸟,j·b·&鸟,M。旅行在南智利和考古学(爱荷华大学出版社,1988年)。
Mayer-Oakes, w·e·塞拉厄瓜多尔北部被网站。反式。点。费罗斯。Soc。76年1 - 14 (1986)。
奈美,h . g .鱼尾抛射点在美洲:言论和假设在南美洲北部的人人。皮疹。Int。578年47 - 72 (2021)。
Waguespack: m & Surovell t·a·克洛维斯狩猎策略,或如何辨认出丰富的资源。点。Antiq。68年(2),333 - 352 (2003)。
Fiedel, s . j .Paleoamerican起源:克洛维斯之外(eds。Bonnichsen, r·莱佩尔b T。,Stanford, D. & Waters M. R.) 97–102 (Texas A&M University Press, 2005).
水域,m . r . &斯塔福德郡,t·w·Paleoamerican奥德赛。在克洛维斯(eds伯爵,k . E。et al。)543 - 562(德州农工大学出版社,2013年)。
林奇,t·F。古印第安人455 - 490(弗里曼,1978)。
g·a·皮尔逊消除距离感:克洛维斯在美国中部的一个更新的概述及其techno-cultural与来自南美的槽点组合。PaleoAmerica3(3),203 - 230 (2017)。
Mackie, m . e . & Haas r估计巧合的频率空间克洛维工件之间的关联和长鼻目动物仍在北美。皮疹。Res。103年,182 - 192 (2021)。
马丁,p . s .美国的发现:第一个美国人可能把西半球,摧毁其在1000年内动物群。科学179年(4077),969 - 974 (1973)。
Surovell, t。水斗式,s R。,Anderson-Sprecher, R. & Myers, A. D. Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna.Proc。国家的。学会科学。美国113年,886 - 891 (2016)。
海恩斯,G。人类世的百科全书1219 - 226 (Springer, 2018)。
格雷森,d . k . & Meltzer d . j .北美过度的安魂曲。j . Archaeol。科学。30.,585 - 593 (2003)。
布劳顿,j . m . & Weitzel e . m .人口重建为人类和巨型建议混合北美更新世灭绝的原因。Commun Nat。91 - 12 (2018)。
Borrero,洛杉矶。美国Megafaunal灭绝的更新世145 - 168 (Springer, 2009)。
马丁内斯,G。,Gutiérrez, M. A. & Tonni, E. P. Paleoenvironments and faunal extinctions: Analysis of the archaeological assemblages at the Paso Otero locality (Argentina) during the Late Pleistocene-Early Holocene.皮疹。Int。299年53 - 63 (2013)。
Miotti, L。,Tonni, E. & Marchionni, L. What happened when the更新世巨型动物灭绝?。皮疹。Int。473年,173 - 189 (2018)。
睡午觉,l·佩雷斯& s i晚更新世南美megafaunal灭绝与鱼尾点和人口有关。Commun Nat。12,2175年。https://doi.org/10.1038/s41467 - 021 - 22506 - 4(2021)。
睡午觉,L。,Politis, G. G. & Perez, S. I. Rapid radiation of humans in South America after the last glacial maximum: A radiocarbon-based study.《公共科学图书馆•综合》15e0236023 (2020)。
皮雷,M。et al。前、中、后megafaunal灭绝:人类影响Pleistocene-Holocene营养网络在巴塔哥尼亚南部。皮疹。科学。牧师。250年106296 (2020)。
Surovell t a &浅滩,b . s .第四纪过度联想的批判和为什么灭绝的争论在很大程度上是无关紧要的。点。Antiq。77年,672 - 687 (2012)。
沃尔夫,A . l . &布劳顿,j . m .觅食理论视角北美更新世过度的联想的批判。j . Archaeol。科学。119年105162 (2020)。
Sisk, m . l . &谢伊,j。j的非洲起源复杂弹技术:分析使用提示横截面积和周长。Int。j .另一个星球。医学杂志。2011年,1 - 8 (2011)。
菲利普斯·J。,Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions.生态。上一次。190年(3 - 4),231 - 259 (2006)。
Guisan,。,Thuiller, W. & Zimmermann, N. E.生境适宜性和分布模型:应用程序在R(剑桥大学出版社,2017)。
Timpson,。et al。重建区域人口波动在欧洲使用放射性碳年代测定新石器时代:一个新的案例研究使用一种改进的方法。j . Archaeol。科学。52,549 - 557 (2014)。
克丽玛,e R。毒蛇,J。,Kobayashi, K. & Madella, M. Summed probability distribution of 14C dates suggests regional divergences in the population dynamics of the Jomon period in eastern Japan.《公共科学图书馆•综合》11(4)、e0154809 (2016)。
布坎南,b &汉密尔顿,m . j .缩放法律Paleoindian弹点的设计。j . Archaeol。方法理论28(2),580 - 602 (2021)。
Sisk, m . l . &谢伊,j。j实验使用性能分析和定量的三角片(Levallois点)作为箭头。j . Archaeol。科学。36(9),2039 - 2047 (2009)。
Dusseldorp, g . l .研究史前狩猎能力:最优觅食理论应用到中间旧石器时代、中石器时代。皮疹。Int。252年3日- 15日(2012年)。
、我。,Bebber, M. R., Knell, E. J., Story, B. & Buchanan, B. Plains Paleoindian projectile point penetration potential.j . Anthropol。Res。78年(1),84 - 112 (2022)。
Friis-Hansen, j .中石器时代的箭头:功能分析的箭头用于狩猎的大型游戏。古代64年(244),494 - 504 (1990)。
Aschero c & Hocsman s . de arqueologia特马。在分析litico(eds Loponte D。et al。)7-26(所de Lujan, 1994)。
布朗,j·L。,Hill, D. J., Dolan, A. M., Carnaval, A. C. & Haywood, A. M. PaleoClim, high spatial resolution paleoclimate surfaces for global land areas.科学。数据。5(1),1 - 9 (2018)。
Sitton, J。,Story, B., Buchanan, B. & Eren, M. I. Tip cross-sectional geometry predicts the penetration depth of stone-tipped projectiles.科学。代表。10(1),1 - 9 (2020)。
卡西尼号,g . H。,Vizcaíno, S. F. & Bargo, M. S. Body mass estimation in Early Miocene native S outh A merican ungulates: A predictive equation based on 3D landmarks.j .黑旋风。287年(1),53 - 64 (2012)。
水域,m R。et al。Pre-Clovis狩猎乳齿象13800年前在客人的网站,华盛顿。科学334年(6054),351 - 353 (2011)。
Waguespack: M。et al。做一个观点:Wood-versus stone-tipped炮弹。古代83年(321),786 - 800 (2009)。
Cione, a . L。,Tonni, E. P. & Soibelzon, L.美国Megafaunal灭绝的更新世125 - 144 (Springer, 2009)。
Tonni e . p . & Carlini a a新第三纪脊椎动物从阿根廷巴塔哥尼亚:他们与最重要的气候变化之间的关系。Dev,皮疹。科学。11,269 - 283 (2008)。
布坎南,B。,Collard, M., Hamilton, M. & O’Brien, M. J. Points and prey: A quantitative test of the hypothesis that prey size influences early Paleoindian projectile point form.j . Archaeol。科学。38,852 - 864 (2011)。
麦特卡尔夫,j·L。et al。协同作用的气候变暖和人类占领在巴塔哥尼亚megafaunal灭绝最后冰消。科学。睡觉。2(6)、e1501682 (2016)。
Moscardi B。,Rindel, D D。&Perez, S. I. Human diet evolution in Patagonia was driven by the expansion of Lama guanicoe after megafaunal extinctions.j . Archaeol。科学。115年105098 (2020)。
Cassiodoro G。,Flores Coni, J., Agnolin, A. & Goñi, R. Caracterización de puntas de proyectil apedunculadas: Un aporte al poblamiento del centro-oeste de la provincia de Santa Cruz (Argentina).航空杂志上亩拉普拉塔。5,126 - 141 (2020)。
苏亚雷斯,r &米洛斯岛的m . j .槽三角non-stemmed点在乌拉圭和extra-regional关系:拓宽技术多样性在全新世早期的南美洲。PaleoAmerica。721 (2021)。
葡方,p .实际Estado de las Investigaciones en el Sitio Arqueologico阿罗约瑞士2(党德非常溢流Provincia de布宜诺斯艾利斯,阿根廷)(ed politi, G。,Scabuzzo, C.) 313–328 (Incuapa-Conicet, 2014).
苏亚雷斯,R。,Piñeiro, G. & Barceló, F. Living on the river edge: The Tigre site (K-87) new data and implications for the initial colonization of the Uruguay River basin.皮疹。Int。473年,242 - 260 (2018)。
Hoguin r &奥克斯曼,b . Palaeoenvironmental场景和岩屑中的第一个人类职业技术阿根廷干高山病。皮疹。Int。363年,78 - 93 (2015)。
布莱恩,a . L。,&Gruhn, R.Taima-Taima的放射性碳年代测定。晚更新世杀死网站Taima-Taima Northermost南部美洲最后1976年发掘的报告(南美第四纪文档计划,密歇根州,1976),页53-58。
jaime, a . e . Vano: El Jobo传统megathere杀死。咕咕叫。更新世》1525日- 27日(1998年)。
Yataco, j . &白痴联赛litica del periodo paleoindio de tipologia joboide, originaria de la半岛de Paraguana al nor-occidente委内瑞拉,en la coleccion del博物馆de Arqueologia y Antropologia de la所市长德圣马科斯。Arqueol。Soc。249-42 (2012)。
Frison, g . c .实验使用克洛维非洲象武器和工具。点。Antiq。54(4),766 - 784 (1989)。
Frison G。生存的狩猎(加州大学出版社,2004年)。
Cruxent, j . m . &唤醒,岩屑被产业类型在委内瑞拉。点。Antiq。22,172 - 179 (1956)。
b .创伤弹道学:木材、j . & Fitzhugh猎物的具体影响渗透从骨的创伤,精疲力竭的石头,和复合插图microblade炮弹在更新世和全新世过渡期间,美国阿拉斯加。j . Archaeol。科学。91年,104 - 117 (2018)。
Grady, J。症结:提示横截面积与周长的有效性识别潜在的岩屑电枢。杜克大学的博士论文(2017)。
休斯,S要点:进化在史前武器。j . Archaeol。方法理论。5(4),345 - 408 (1998)。
Weitzel C。,Flegenheimer, N., Martinez, J. & Colombo, M. Breakage patterns on fishtail projectile points: Experimental and archaeological cases.种族文化考古学。6(2),81 - 102 (2014)。
苏亚雷斯,r .评论南美鱼尾点:设计、序列,减少和功能。咕咕叫。更新世》23,78 - 80 (2006)。
Carbonera, m & Loponte d .原材料和鱼尾弹点从巴西南部的功能设计。j .岩屑钉。81 - (2021)。
丘吉尔,s e .武器技术,猎物的大小选择,对狩猎和狩猎方法在现代狩猎采集者:影响在旧石器时代、中石器时代。Archeol。人民行动党。点。Anthropol。Assoc。4(1)11-24 (1993)。
、我。et al。克洛维斯的功效的槽点狩猎长鼻目动物。J Archaeol。科学。39103166 (2021)。
油渣,r D。弹技术287 - 320 (Springer, 1997)。
Hocsman, S。,Martínez, J. G., Aschero, C. A., & Calisaya, A. in南行:拉丁美洲的更新世后期工作(Eds。Miotti, L。,Salemme, M. Flegenheimer, N. & Goebel, T.) 63–68 (Texas A&M University Press, 2012).
Hoguin, R。,&Restifo, F. in南行:拉丁美洲的更新世后期工作(Eds。Miotti, L。,Salemme, M. Flegenheimer, N. & Goebel, T.) 69–74 (Texas A&M University Press, 2012).
苏亚雷斯,R。Arqueologia杜兰特拉跃迁Pleistoceno-Holoceno en乌拉圭。国际系列2220酒吧(牛轭出版社,2011年)。
海德,g . & Rivero d工厂化morfometricos aplicados制盐业de proyectil lanceoladas del Holoceno temprano-medio en内华达山脉y llanuras坐落de阿根廷。纬度。点。Antiq。29日(3),572 - 590 (2018)。
万利拉,l . &淀粉,r . a .同现mylodontid树懒和见解的潜在分布在晚更新世。皮疹。Res。85年(1),66 - 74 (2016)。
Rindel, D D。,Moscardi B。F. & Perez, S. I. The distribution of the guanaco (Lama guanicoe) in Patagonia during Late Pleistocene-Holocene and its importance for prehistoric human diet.全新世31日(4),644 - 657 (2021)。
QGIS开发团队。QGIS地理信息系统。开源项目地理空间基础。http://qgis.osgeo.org(2021)。
r开发核心团队。接待员:统计计算的语言和环境。(R统计计算的基础,2021)。http://www.R-project.org/。
Villavicencio, N。晚第四纪Megafaunal灭绝在南美:年表、环境变化和人类影响区域尺度。博士论文(加州大学2016)。
确认
我们感谢斯图尔特Fiedel,迭戈Rindel Matias麦地那的有益的建议和评论初步版本的手稿,塞萨尔门德斯,Luis Borrero Gustavo马丁内斯,诺拉Flegenheimer,所罗门Hocsman,雨果Nami,费德里科•Restifo,拉斐尔·苏亚雷斯和古斯塔沃politi提供参考书目和未发表的数据。
资金
所提供的资金是de La Plata(授予11号/ N885, 11 / N932), Conicet Trianual格兰特(赠款。PIP3204和PIP2974)和ANPCyT Trianual批准(批准号2019 - 04547)。
作者信息
作者和联系
贡献
概念化:石油醚和S.I.P.方法:湄S.I.P.调查:石油醚,是由,S.I.P. Visualization: S.I.P. Writing—original draft and editing: L.P.
相应的作者
道德声明
相互竞争的利益
作者宣称没有利益冲突。
额外的信息
出版商的注意
施普林格自然保持中立在发表关于司法主权地图和所属机构。
权利和权限
开放获取本文是基于知识共享署名4.0国际许可,允许使用、共享、适应、分布和繁殖在任何媒介或格式,只要你给予适当的信贷原始作者(年代)和来源,提供一个链接到创作共用许可证,并指出如果变化。本文中的图片或其他第三方材料都包含在本文的创作共用许可证,除非另有说明在一个信用额度的材料。如果材料不包括在本文的创作共用许可证和用途是不允许按法定规定或超过允许的使用,您将需要获得直接从版权所有者的许可。查看本许可证的副本,访问http://creativecommons.org/licenses/by/4.0/。
关于这篇文章

引用这篇文章
睡午觉,L。,Rivero, D. & Perez, S.I. Changes in projectile design and size of prey reveal the central role of Fishtail points in megafauna hunting in South America.Sci代表1216964 (2022)。https://doi.org/10.1038/s41598 - 022 - 21287 - 0
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s41598 - 022 - 21287 - 0
