







摘要
扩展金丝桃科植物的基因组信息,特别是在Cratoxylum,我们鉴定了5个新的完整质体基因组(质体组)Cratoxylum和它的两个同属分类群,包括c . arborescens,c . formosum无性系种群。formosum,c . formosum无性系种群。pruniflorum,c . maingayi,c . sumatranum,金丝桃属植物hookerianum,Triadenum breviflorum。为Cratoxylum质粒长度在156,962 ~ 157,792 bp之间。5个质体的基因组结构和基因含量由128-129个基因组成,其中包括83-84个蛋白质编码(CDS)基因、37个tRNA基因和8个rRNA基因。的质体h . hookerianum和t . breviflorum分别为138,260 bp和167,693 bp。共有110个和127个基因,包括72个和82个CDS, 34个和37个tRNA, 4个和8个rRNA基因。利用最大似然树(ML)和贝叶斯推理树(BI)重建系统发育树,发现在属水平上具有相同的拓扑结构;Cratoxylum是单元。然而,c . formosum无性系种群。pruniflorum没有和它的起源聚集在一起,这让人怀疑它是否应该被视为一个独立的物种,c . pruniflorum基于形态学描述支持的分子证据。
介绍
金丝桃科在世界范围内共有9属500余种。一般来说,金丝桃科的成员可以进一步分为三个不同的部落,即:克拉托克叶(Cratoxyleae Bentham & J.D. Hooker)、金丝桃科(Hypericeae Choisy)和金丝桃科(Vismieae Choicy)1。作为家族中最小的部落,两个属被认为是在Cratoxyleae,即。CratoxylumBlume和单型属,ElieaCambess2。目前,有7种被认可的Cratoxylum布卢姆(金丝桃科,malpiighiales),其中三个被认为具有至少两个种内身份3.。的成员Cratoxylum原产于亚洲热带地区,从印度到中国南部再到马来西亚分布广泛,人们经常寻找它们的木材作为木材和木炭生产的来源4。这些物种对恶劣环境的良好适应性和快速生长的性能使它们成为潜在的补种物种,对泥炭地的恢复策略有用5,6。
尽管在泥炭沼泽森林中有作为有用的恢复剂的潜力,遗传研究Cratoxylum是有限的。遗传数据Cratoxylum是否只局限于来自质体、线粒体和核区的短基因序列C。arborescens(嗯)好吧c . cochinchinense不悦之色。作为其属的代表种的枫香属植物系统发育树的重建7,8,9。物种间系统发育研究的缺乏Cratoxylum阻碍了我们在遗传水平上对这个属的理解。
质体基因组是分子分类学研究的宝贵资源。被子植物质体是由一个大单拷贝(LSC)区、两个反向重复(IR)区和一个小单拷贝(SSC)区组成的圆形单倍体基因组,其长度通常在110-240 kbp之间10。近年来,研究人员对利用新一代测序技术获得完整的质体序列表现出极大的兴趣。这是因为与短基因序列相比,用于系统发育的基因组尺度数据集包含更多的单核苷酸多态性,这有助于重建一个得到良好支持的系统发育树11。由于测序技术的进步和有用的生物信息学程序的可用性来帮助组装和注释质体体,到目前为止,许多完整的质体体序列已经公开,用于解释复杂属中模糊的系统发育关系12,13。另一方面,核DNA内部转录间隔区(ITS)在核基因组水平上揭示植物的双亲本遗传是有用的14。在所有的核基因中,ITS序列的扩增是已知的更容易的,并且所提供的遗传信息也有助于在种内水平上划分个体15。
尽管近年来已发表的完整质体的记录大量增加,但金丝桃科的基因组数据仍然缺乏。到目前为止,已发表的记录的完整的质体序列Cratoxylum仅限于c . cochinchinense,其中NCBI GenBank数据库中至少有三个不同加入的基因组序列(截至2021年9月)。为了进一步丰富金丝桃科植物的基因组信息,我们对金丝桃属5个分类群的质体体进行了进一步的测序和鉴定Cratoxylum,包括c . arborescens,c . formosum无性系种群。formosum(杰克)Benth。& Hook.f。戴尔,c . formosum无性系种群。pruniflorumKurz Gogelein,c . maingayi戴尔,c . sumatranum(杰克)布卢姆,以及两个近亲物种,金丝桃属植物hookerianumwright & Arn。和Triadenum breviflorum墙。前戴尔。为了揭示植物种间的系统发育关系Cratoxylum研究人员进一步利用质体蛋白编码序列(CDS)数据集和细胞核DNA内部转录间隔区(ITS)序列区进行了系统发育分析。本研究结果将为金丝桃科和麻瓜属植物的系统发育和进化研究提供重要参考。
结果与讨论
质体系特点
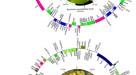
从这项研究中获得的所有七个质体都表现出典型的四分体结构,由一个大的单拷贝(LSC)和一个小的单拷贝(SSC)区域组成,它们被一对反向重复序列(IR)分开(图2)。1).质体体大小在156,962 bp (c . formosum无性系种群。pruniflorum157,792 bp (c . arborescens)是属的五个分类群之一Cratoxylum,而h . hookerianum和t . breviflorum长度分别为138,260 bp和167,693 bp(表1).在5个分类群的质体体中共预测到128 ~ 129个基因Cratoxylum该基因由83-84个CDS、37个tRNA和8个rRNA基因组成。Cratoxylum formosum无性系种群。formosum和c . formosum无性系种群。pruniflorum与其他三种相比,我们缺少一种CDSCratoxylum,这是rpl32个基因应位于SSC区(表2)2).有15个基因,包括9个CDS和6个tRNA基因,包含一个内含子,而2个基因,中电控股P和ycf3,含有两个内含子。

本研究所用金丝桃科7个分类群的基因组结构和基因图谱。Cratoxylum arborescens,c . maingayi,c . sumatranum(一个);Cratoxylum formosum无性系种群。formosum,c . formosum无性系种群。pruniflorum(B);金丝桃属植物hookerianum(C);Triadenum breviflorum(D).内圈基因顺时针转录,外圈基因逆时针转录。颜色代码描述了基因的不同功能组。粗线表示反向重复序列(IRA和IRB)的边界,在大单拷贝(LSC)和小单拷贝(SSC)区域之间划分。内圈深灰色区域为基因组GC含量,浅灰色区域为AT含量。
虽然在质体中的基因含量Cratoxylum在本研究中检测的5个分类群中是一致的,在LSC区域检测到一个反向的基因块排列,在加拿大皇家银行L和的环境K-UUU基因(图1).逆转录基因块长度约为55000 bp,包含28个CDS和19个tRNA基因。通过与其他近缘家族的质体进行比较,我们发现该基因的基因排列阻滞在c . cochinchinense,c . formosum无性系种群。formosum,c . formosum无性系种群。pruniflorum与龙舌兰科、龙舌兰科、金舌兰科和龙舌兰科相似;Bonnetia香(GenBank登记号:MK995182),Caraipa heterocarpa(GenBank登记号:MW853787),藤黄属植物mangostana(GenBank登记号:KX822787),Licania micrantha(GenBank登记号:KX180080);而基因的排列被阻断了c . arborescens,c . maingayi,c . sumatrana与菊科植物相同,即Marathrum capillaceum(GenBank登记号:MN165813)和藻trifaria(GenBank登记号:MK995179)。这一发现与之前的一项研究一致,在该研究中,基因块反转在加拿大皇家银行L和acc在金丝桃科(Hypericaceae)、足草科(Podostemaceae)和凤蝶科(Papilionoideae)两个拟合科中均发现D16。为h . hookerianum和t . breviflorum,共预测到110个和127个基因,其中CDS分别为72个和82个,tRNA分别为34个和37个,rRNA分别为4个和8个。测定了5个类群的质体GC含量Cratoxylum色谱含量在36.1% ~ 36.3%之间h . hookerianum和t . breviflorum分别为38.1%和37.4%。
短序列和大序列重复
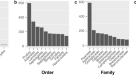
简单序列重复序列(SSRs)或微卫星是1-6个核苷酸和特定位点基序的短串联重复序列,存在于所有基因组中,特别是真核生物。除了作为基因组标记用于标记辅助选择、亲缘关系、育种等方面外,SSRs还通过其编码区长度的变化发挥重要的调控功能17,18。在本研究中,检测到的总SSRsc . arborescens,c . cochinchinense,c . formosum无性系种群。formosum,c . formosum无性系种群。pruniflorum,c . maingayi,c . sumatranum分别为170、103、95、95、104、96。2).在所有重复类型中,单核苷酸重复最为丰富,在69 (c . formosum无性系种群。formosum)及80 (c . arborescens);A/T型单核苷酸重复频率大于C/G型重复频率。值得注意的是,五核苷酸只存在于c . arborescens,包括两种AAATT/AATTT和一种AAAAT/ attt重复类型。在评估的六个质体中,大的重复仅以正向重复和回文重复的形式记录。所有的质粒都被鉴定为25个,除了c . sumatranum它有24个正向重复和26个回文重复。

本研究使用的金丝桃科7个分类群的质体基因组重复序列。不同简单序列重复类型数量(一个);分类SSR重复单位的数目(B);长重复(包括正向重复和回文重复)的分布和频率(C).
红外区域的膨胀和收缩
6个分类群的质质体IR连接邻近基因Cratoxylum所检测的基因含量相同(图2)。3.).连接LSC/IRb (JLB)的邻近基因为rpl22日,石头剪刀19日,石头剪刀19个位于JLB。然而,对于LSC/IRa结(JLA),石头剪刀爱尔兰共和军地区有19人完好无损,而的环境记录到LSC区域的H穿过JLA。的ycf1个基因被放置在所有6个类群的SSC/IRa交界处(JSA)Cratoxylum但对于接合SSC/IRb (JSB),只有ycf四个分类群的1个基因Cratoxylum,包括c . cochinchinense,c . formosum无性系种群。formosum,c . maingayi,c . sumatranum,被发现放置在JSB上。的ycf的第1c . arborescens在IRb区域仍然完好无损,而ycf1基因c . formosum无性系种群。pruniflorum被鉴定为长度较短,推测为假基因,位于SSC区。与植物的6个分类群一起分析Cratoxylum,与IR连接点相邻的基因含量h . hookerianum和t . breviflorum表现出一些变化,当比较Cratoxylum。在JLA和JLB,基因含量t . breviflorum是否与…相似Cratoxylum,其中石头剪刀19岁,的环境H分别放置在JLB和JLA之间。然而,对于h . hookerianum,的环境N基因放置在LSC区,JLB旁边加拿大皇家银行LSC区的L基因与JLA的距离最近。这两个h . hookerianum和T。breviflorum有ndhF基因位于IR区,与JSA和JSB相邻,而位于SSC区;ndh一个和石头剪刀15个被放置在JSA旁边h . hookerianum和t . breviflorum,分别。

基于金丝桃科8个分类群全质体基因组的IR边界分析c . arborescens,c . cochinchinense,c . formosum无性系种群。formosum,c . formosum无性系种群。pruniflorum,c . maingayi,c . sumatranum,金丝桃属植物hookerianum,Triadenum breviflorum。
比较基因组分析
全质体组序列的基因组比较分析显示,该物种在所有类群中均具有高度保守性Cratoxylum,c . cochinchinense作为参考基因组(图2)。4).在基因间隔区观察到一个类似于缺失形式变异的小间隙公安局J -宠物一个的c . formosumsubp。formosum,c . formosum无性系种群。pruniflorum,c . sumatranum。当与h . hookerianum和t . breviflorum,在质体上至少可以观察到6个大的间隙,这表明在属水平上核苷酸序列有很大的差异。这些缺口位于的环境Q-UUG -的环境K-UUU,中电控股P,的环境R-ACG -ndhF,的环境B-ACG -石头剪刀15和2ycf2区域。植物6个分类群的多序列比对Cratoxylum全长169,031 bp,包含1447个单点和22,090个简约信息位点。至少有1678个indel事件被确定,包括23,364个indel站点。

本研究使用的金丝桃科7个分类群的基因组比较分析,获得了金丝桃科的全质体基因组序列Cratoxylum cochinchinense(MN399961)作为参考基因组。在Shuffle-LAGAN模式下使用mVISTA进行分析。图例用颜色编码描述了基因区域的方向和类型。概率阈值设为50%。黑色箭头表示在质体基因组中显示明显分化的区域。
系统发育推断
拼接后的CDS数据集的多序列比对总长度为89,82 bp,而ITS数据集的多序列比对在修剪前和修剪后分别为807 bp和723 bp。由于ML树和BI树在两个数据集中都具有相同的拓扑结构,因此本研究只呈现ML树,并在每个分支节点上包含BI后验概率。在cds树中,所有分支节点均得到良好支持(BS:≥75;pp:≥0.95);研究中所有分类群之间的系统发育关系得到了很好的解决(图2)。5一个)。Cratoxylum结果表明,6个类群均为单系关系。Cratoxylum arborescens首先从其他分类群中分化出来,其次是c . cochinchinense,c . formosum无性系种群。pruniflorum,c . maingayi。Cratoxylum formosum无性系种群。formosum被放在树枝的顶端c . sumatranum。对于ITS数据集,与使用cds数据集重建的树相比,观察到类似的树拓扑结构(图2)。5B);单系关系也存在于Cratoxylum,其中所有六个分类群的分子位置Cratoxylum在its树中显示的与在cds树中显示的相同。植物各类群的系统发育关系Cratoxylum在使用ITS数据集(BS:≥75;pp:≥0.95)。

的系统发育树Cratoxylum以及基于来自质体基因组的串联蛋白质编码序列的相关分类群(一个),以及核DNA内部转录间隔序列(ITS) (B).利用最大似然(ML)和贝叶斯推理(BI)构建了系统发育树。Bootstrap支持(BS)和后验概率(PP)被认为是可靠的(BS:≥75;PP:≥0.95)用星号(*)表示。
关于植物分类身份的冲突c . formosum无性系种群。Pruniflorum
值得注意的是c . formosum无性系种群。pruniflorum没有和原来的组合在一起,c . formosum无性系种群。formosum在用核和质体区重建的系统发育树中。尽管ITS序列是双亲本遗传,但可能表明杂交c . formosum无性系种群。pruniflorum;然而,基于母体遗传质体基因的系统发育树显示c . formosum无性系种群。formosum和c . formosum无性系种群。pruniflorum应该作为两个自然的群体来对待。根据文献,c . formosum无性系种群。pruniflorum最初被认为是一个独特的物种,金丝桃属植物prunifolium墙19。然而,该物种经历了几次分类修订,才被认为是一个亚种c . formosum1967年4。在描述的基础上,作者强调了区分的关键c . formosum无性系种群。pruniflorum它的起源是基于一份文件的出现;两个类群表现出高度的形态相似性。虽然这不是区分两个分类群的关键形态学特征,c . formosum无性系种群。pruniflorum带有短柔毛萼片,而c . formosum无性系种群。formosum所有部分都无毛,它们在地理上是明确的,几乎没有重叠。其他形态学特征被提议用来划定界限c . formosum无性系种群。pruniflorum从c . formosum无性系种群。formosum前者有生锈的,被绒毛的幼枝,花梗和花萼,而后者无毛;前者为短而截形的下鳞,长0.7-0.8 mm,后者为舌形的下鳞,长2 mm;前者蒴果为卵形,内含54-58粒种子;后者蒴果为椭球状,内含36-46粒种子20.。
以确定遗传距离之间c . formosum无性系种群。pruniflorum基于完整质体体和its序列,对完整质体体和its序列进行两两距离分析,分别进行分析。我们发现种内两两距离为0.00351,大于种间两两距离c . formosum无性系种群。pruniflorum和c . maingayi在质体水平上(0.00159),种内两两距离为0.0502,大于种间两两距离c . formosumsubp。pruniflorum和c . maingayi(0.0358)以及c . sumatranum(0.0316)在ITS水平。没有自然杂交的报道Cratoxylum;尽管两两距离不是区分近缘种的合适参数,但通常认为种内两两距离应小于种间两两距离21,22。另一方面,染色体计数c . cochinchinensen = 1123,而c . formosum无性系种群。formosumn = 724。一般来说,在自然环境下,二倍体植物种内的染色体数往往是保守的25。Cratoxylum cochinchinense被提议与c . formosum一度因其形态特征相同,但该提议后来被否决;在树的大小、树皮的颜色、叶的结构、叶的形状和花的雄蕊束等方面,两种植物的形态差异明显4。因此,我们评论细胞学研究c . formosum无性系种群。pruniflorum与原始亚种相比,可以提供有用的遗传特性见解。然而,传统的分类方法和分子系统发育分析之间的发现Cratoxylum部分不一致;的分类特性c . formosum无性系种群。pruniflorum被接受为一个退化的分类单元c . formosum对该属作进一步的分类学修订。基于本研究的分子证据,我们认为c . formosum无性系种群。pruniflorum应视为自然类群,而物种名称呢Cratoxylum pruniflorum库尔兹应该复职。
结论
本研究扩大了金丝桃科5个分类群的基因组数据Cratoxylum,以及来自金丝桃属植物和Triadenum。利用CDS和ITS数据集重建的系统发育树得到了很好的解决方案,这一发现为研究植物的分子定位和进化提供了新的思路Cratoxylum,其中的分类特性c . formosum无性系种群。pruniflorum被认为是亚种c . formosum是有问题的。尽管如此,本研究获得的分子数据将为更好地了解金丝桃科植物的分类和系统发育提供宝贵的资源。
材料与方法
植物材料
五个植物分类群的新鲜叶子Cratoxylum物种,包括c . arborescens,c . formosum无性系种群。formosum,c . formosum无性系种群。pruniflorum,c . maingayi,c . sumatranum,以及……金丝桃属植物hookerianum和Triadenum breviflorum(补充图。1)从自然种群和迁地地点收集(表1)1).所有实验均按照相关指南和规定进行。每个标本的身份在标本采集之前由相应的作者确认。叶片标本保存在装满硅胶的自封袋中,运送到各自的当地实验室进行总基因组DNA提取。
DNA提取,基因组测序和组装
根据制造商的方案,使用DNeasy Plant Mini Kit (QIAGEN, Germany)进行总基因组DNA检测。使用Qubit™4荧光仪(Thermo Fisher Scientific, USA)估计DNA提取物的纯度和数量。下一代测序在Illumina NovaSeq平台(Illumina, USA)上进行,获得350-bp的配对末端reads。使用NGS QC Toolkit v2.3修剪适配器序列26使用NOVOPlasty v2.7.2组装质体27与加拿大皇家银行L基因c . cochinchinense(GenBank登记号:MN399961)作为种子序列。用GeSeq v2.03对组装的质体进行注释,并对倒置区连接进行鉴定28。手动检查已注释的质体是否有错误。使用OGDRAW v1.3.1可视化圆形质体图29。本研究获得的所有质体组序列均存入NCBI GenBank数据库,登录号为MZ703415-MZ703419, MZ714015-MZ714016。
重复分析
为了提供一个更好的了解质体之间的所有物种的Cratoxylum可在线获取,完整的质体序列c . cochinchinense(GenBank登记号:MN399961)从NCBI GenBank数据库下载。随后的基因组比较分析纳入了该物种的基因组数据。利用MISA-web,鉴定了每个质体的ssr30.。对单核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸基序分别设置最小重复参数数为10、4、4、3、3和3。大的重复序列,包括正向重复序列、回文重复序列、反向重复序列和补体重复序列,使用repter进行鉴定31,其中最小重复大小为30 bp,汉明距离为3。
基因组比较和序列差异分析
为了检测质体中红外区域的扩张和收缩,使用IRscope程序对红外区域的边界和连接处进行了可视化32并使用Adobe Photoshop CS6 (Adobe, USA)进行进一步编辑。使用mVISTA进行基因组比较分析33在Shuffle-LAGAN模式下进行基因组比对。的质体序列c . cochinchinense(GenBank登记号:选择MN399961)作为参考基因组。使用MAFFT v7进行的多基因组比对中存在的多态性、简约性信息和缺失位点的数量34也使用DnaSP v5.10.0135。
聚合酶链反应和桑格测序
获得了中国野生植物5个分类群的ITS (ITS1-5.8S-ITS2)序列Cratoxylum,以及……h . hookerianum,t . breviflorum本研究采用一对通用引物its55′-GGAAGTAAAAGTCGTAACAAGG-3′和its45′-TCCTCCGCTTATTGATATGC-3′进行聚合酶链反应(PCR)。36。最终反应体积为25µL,加入12.5µL的2 × GoTaq,进行PCR扩增®Green Master Mix (Promega, USA),正向和反向引物各10µM, DNA模板15 ng。PCR扩增与热设置一个初始编程变性在95°C 2分钟,其次是30周期在95°C的变性30年代,退火55°C 30年代,在72°C扩展1分钟,最后在72°C扩展5分钟。扩增子通过凝胶电泳验证和查看下UV机之前发送直接Sanger测序两端使用ABI 3730 DNA分析仪(美国应用生物系统公司)。使用MEGA7对结果序列进行对齐和手动编辑37获得将用于系统发育分析的干净序列。本研究获得的ITS序列已存入NCBI GenBank数据库,登录号为MZ674200-MZ674204、MZ703053和OM980718。
系统发育重建
利用14个分类群的CDS序列进行了基于CDS的系统发育树重建,其中金丝桃科8个,近缘种7个。Bonnetia香(金丝桃科;GenBank登记号MK995182),Caraipa heterocarpa(Calophyllaceae;GenBank登记号MW853787),藤黄属植物mangostana(金丝桃科;GenBank登记号KX822787),Licania micrantha(可可李科;GenBank登记号KX180080),Marathrum capillaceum(河苔草科;GenBank登记号MN165813),Mesua ferrea(Calophyllaceae;GenBank登记号NC_049111),以及藻trifaria(河苔草科;GenBank登记号MK995179)属于Malpighiales一起分析。Averrhoa杨桃(酢浆草料;GenBank登记号将Oxalidales的KX364202作为外群。利用MAFFT v7对质体体序列进行比对34采用最大似然(ML)和贝叶斯推理(BI)方法进行系统发育分析。在ML分析中,建立了一个具有gamma (+ G) (= GTR + G)的广义时间可逆(GTR)模型,并使用RAxML v8.2.11在1000次bootstrap重复下重建了ML树38。使用MrBayes v3.2.7a进行BI分析39在CIPRES科学网关中提供的管道40。似然模型选择混合替代型和4 × 4核苷酸替代模型,实现200万代马尔可夫链蒙特卡罗分析和4条马尔可夫链。每100代进行一次数据采样,而前25%的树木被作为烧毁而丢弃。使用FigTree v.1.4.4将两种分析的最终树结果可视化41。
基于ITS序列重建了棘叶属7个分类群的系统发育树,其中包括棘叶属5个分类群Cratoxylum在这项研究中,c . cochinchinense(GenBank登记号:KP092947),Eliea关节动物(GenBank登记号:KC709409)。金丝桃属植物hookerianum和t . breviflorum被列为外群体。利用MEGA7中嵌入的MUSCLE进行多序列比对36使用trimAL v1.2对线进行修整42通过选择间隙输出选项来减少由于对准不良而产生的系统误差。使用MEGA7进行ML分析37,其中MEGA7中嵌入的“Find Best DNA/Protein Model (ML)”功能计算出包含(+ I)不变位点的Tamura 3-parameter (T92)模型(= T92 + I)是最优的DNA替代模型。所有位点均纳入分析,并以1000个bootstrap重复进行计算。对于BI分析,使用MrBayes v3.2.7a进行计算39遵循与上面提到的相同的参数和设置。
数据可用性
支持本研究结果的数据可在NCBI的GenBank中公开获取https://www.ncbi.nlm.nih.gov,登录号(MZ674200-MZ674204, MZ703053, MZ703415-MZ703419, MZ714015-MZ714016, OM980718)。支持本研究结果的原始NGS数据可应通讯作者a.c.的合理要求提供。
参考文献
被子植物系统发育网站。14版本。http://www.mobot.org/MOBOT/research/APweb(2017)。
史蒂文斯,p.f.金丝桃科。在开花植物·Eudicots(克劳斯主编,K.) 194-201 (b施普林格,2007)。
POWO。世界植物在线。由邱园皇家植物园协助。https://powo.science.kew.org(2022)。
葛葛林,A. J. F.植物属的修订Cratoxylum提单(藤黄科)。艾纳香属15, 453-475 (1967)
庄新。香港退化丘陵上森林的恢复与发展。对。生态。等内容。99, 1997 - 2011(1997)。
Suwito, D., Suratman, & Poedjirahajoe, E.Cratoxylum arborescensBlume (Geronggang)和Combrecarpus rotundatus丹斯(Tumih)在退化的热带泥炭沼泽森林中作为自然再生。JISDeP2, 272-289(2021)。
Tokuoka, T. & Tobe, H.使用质体和核DNA序列对malpiighiales进行系统发育分析,特别参考了大戟科植物的胚胎学。植物研究。119, 599-616(2006)。
Wurdack, K. J. & Davis, C. C. malpiighiales的系统发育:在被子植物生命树上最顽固的分支之一上取得进展。点。j .机器人。96, 1551-1570(2009)。
鲁赫费尔,b.r.。et al。闭尾猿进化支的系统发育:来自质体和线粒体基因组的证据。点。j .机器人。98, 306-325(2011)。
刘建军,张建军,张建军,张建军,张建军。植物叶绿体染色体的进化:基因含量、基因序列和基因功能。植物分子生物学76, 273-297(2011)。
Moore, M. J., Soltis, P. S., Bell, C. D., Burleigh, J. G.和Soltis, D. E.对83个质体基因的系统发育分析进一步解决了早期分化的问题。Proc。国家的。学会科学。美国107, 4623-4628(2010)。
霁,Y。et al。质体体系统基因组学、生物地理学和进化枝多样化巴黎(Melanthiaceae)。BMC植物生物学。19, 543年。https://doi.org/10.1186/s12870-019-2147-6(2019)。
Mehmood, F., Rahim, A., Heidari, P., Ahmed, I. & Poczai, P.比较质体分析艾纳香属,对小行星科的基因组进化和系统发育具有启示意义。生态。另一个星球。11, 7810-7826(2021)。
植物DNA序列的系统发育分析:从植物到序列。方法Mol. Biol;718, 183-235(2011)。
Álvarez, I. J. F. W. &温德尔,J. F.核糖体ITS序列与植物系统发育推断。摩尔。Phylogenet。另一个星球。29, 417-434(2003)。
金东明,金建军,易天生,等。马尔皮亚目拟龙枝质体结构保护与进化。科学。代表。10, 9091年。https://doi.org/10.1038/s41598-020-66024-7(2020)。
gie, s.h., Das, p.u s.d.和Sharma, m.p.用DNA标记鉴定药用植物。植物基因4, 83-89(2015)。
Avvaru, A. K, Saxena, S., Sowpati, D. t和Mishra, R. K. MSDB:一个简单序列重复的综合数据库。基因组医学杂志。另一个星球。9, 1797-1802(2017)。
第7276号金丝桃属植物prunifolium。号码。列表[Wallich]。245(1832)。
S. N.比斯瓦斯Cratoxylumformosumssp。pruniflorumKurz高格。(金丝桃科)。莲属15, 167-169(1973)。
Meier, R., Shiyang, K., Vaidya, G.和Ng, P. K.双翅目这是一个种内变异性高、鉴定成功率低的故事。系统。医学杂志。55, 715-728(2006)。
周玉芳。et al。基因流动与物种划分——以中国东南部重叠分布的两种松树为例。另一个星球。Int。j . Org。另一个星球。64科学通报,2342-2352(2010)。
米歇尔,C。Cratoxylum(PROSEA)。植物使用英语。https://uses.plantnet-project.org/e/index.php?title=Cratoxylum_ (PROSEA) &oldid = 327154(2017)。
N. K. B.罗布森植物区系金丝桃科第8卷,1-29 (Alpha版,1974年)。
蔡志强,张涛,简惠英,一种有发展前景的油料木本作物染色体数目变异,Plukenetia瓦卢l .(大戟科)。Caryologia66, 54-58(2013)。
帕特尔,r.k.和贾恩,M. NGS质量控制工具包:下一代测序数据质量控制工具包。《公共科学图书馆•综合》7, e30619。https://doi.org/10.1371/journal.pone.0030619(2012)。
Dierckxsens, N., Mardulyn, P. & Smits, G. NOVOPlasty:从全基因组数据中重新组装细胞器基因组。核酸类。45, e18。https://doi.org/10.1093/nar/gkw955(2017)。
蒂利希,M。et al。geseq -细胞器基因组的通用和准确注释。核酸类。45, w6-w11(2017)。
Greiner, S., Lehwark, P. & Bock, R. Organellar Genome DRAW (OGDRAW) 1.3.1版:扩展了Organellar基因组的图形可视化工具包。核酸类。47, w59-w64(2019)。
Beier, S., Thiel, T., m
库尔茨,S。et al。重复序列分析在基因组尺度上的多种应用。核酸类。29(2001)。
Amiryousefi, A., Hyvӧnen, J. & Poczai, P. IRscope:叶绿体基因组连接位点可视化的在线程序。生物信息学34, 3030-3031(2018)。
弗雷泽,k.a.,帕切特,L.,波利亚科夫,A.,鲁宾,e.m.和杜布切克,I. VISTA:比较基因组学的计算工具。核酸类。32, w273-w279(2004)。
kaoh, K, Rozewicki, J. & Yamada, K. D. MAFFT在线服务:多序列比对、交互式序列选择和可视化。简短。Bioinform。20.中文信息学报,1160-1166(2019)。
Librado, P. & Rozas, J. DNA多态性数据综合分析软件。生物信息学25, 1451-1452(2009)。
白,T. J,布朗,T. D,李,S. b和泰勒,J. W.真菌核糖体RNA基因的系统发育扩增和直接测序。在PCR协议:方法和应用指南英尼斯,文学硕士。et al。315-322(学术出版社,1990)。
Kumar, S., Stecher, G., Li, M., Knyaz, C.和Tamura, K. MEGA X:跨计算平台的分子进化遗传学分析。摩尔。杂志。另一个星球。35中文信息学报,1547-1549(2018)。
RAxML version 8:用于系统发育分析和大型系统发育后分析的工具。生物信息学30.科学通报,1312-1313(2014)。
Ronquist, F, Teslenko, M, Mark, P. V. d和Huelsenbeck, J. mr . bayes 3.2:跨大模型空间的高效贝叶斯系统发育推断和模型选择。系统。医学杂志。61, 539-542(2012)。
Miller, m.a, Pfeiffer, W. & Schwartz, T. CIPRES科学门户:系统发育分析的社区资源。在2011年TeraGrid会议论文集:极限数字发现41.https://doi.org/10.1145/2016741.2016785(2011)
A.兰博特。http://tree.bio.ed.ac.uk(2018)。
capella - gutisamrrez, S, Silla-Martinez, J. M. & Gabaldon, T. TrimAl:一种大规模系统发育分析中的自动对齐修剪工具。生物信息学25, 1972-1973(2009)。
致谢
这项工作得到了孔敬大学研究与研究生院的支持。作者感谢西南野生物种种质资源库为其提供的DNA材料Triadenum breviflorum在这项研究中。
作者信息
作者及单位
贡献
概念化,S.Y.L, A.C.和R.S.;方法论,r.s.,美国;形式分析,r.s., s.k., S.Y.L.;资源,P.S.和S.A.S.;数据管理,p.s., S.Y.L.;写作-原稿准备,r.s., S.K.;写作、评审和编辑,s.a.s., s.y.l., A.C.;监督、通讯、交流;所有作者已阅读并同意稿件的出版版本。
相应的作者
道德声明
相互竞争的利益
作者声明没有利益冲突。
额外的信息
出版商的注意
b施普林格《自然》杂志对已出版的地图和机构的管辖权要求保持中立。
补充信息
权利和权限
开放获取本文遵循知识共享署名4.0国际许可协议,该协议允许以任何媒介或格式使用、共享、改编、分发和复制,只要您适当地注明原作者和来源,提供知识共享许可协议的链接,并注明是否进行了更改。本文中的图像或其他第三方材料包含在文章的知识共享许可协议中,除非在材料的署名中另有说明。如果材料未包含在文章的知识共享许可中,并且您的预期用途不被法律法规允许或超过允许的用途,您将需要直接获得版权所有者的许可。如欲查阅本许可证副本,请浏览http://creativecommons.org/licenses/by/4.0/。
关于本文

引用本文
Sudmoon, R., Kaewdaungdee, S., Tanee, T.;et al。植物质体基因组的特征Cratoxylum种(金丝桃科)和系统发育关系的新见解。Sci代表12, 18810(2022)。https://doi.org/10.1038/s41598-022-23639-2
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s41598-022-23639-2
