








摘要
三叶虫是典型的古生代生物矿化海洋真节肢动物,在寒武纪“大爆发”期间突然出现在化石记录中(约5.21亿年前)。自从达尔文对地球上缺乏前三叶生物化石感到困惑以来,这种突然的出现就一直备受争议物种起源一般认为,三叶虫一定有一个未被观察到的神秘的进化史,可以追溯到前寒武纪。在这里,我们回顾了这个模型背后的假设,并提出了一个神秘的历史造成了重大的困难,包括调用生物矿化结构的疯狂趋同进化和放弃联合进化支的突触形态。我们发现,早寒武纪三叶虫古地理格局的地理分隔解释与控制现存海洋无脊椎动物分布的因素不一致,包括越来越多的人认识到长距离传播的重要性。我们认为生存偏差可能解释了三叶虫最初的快速多样化,并得出结论,该类群在约521 Ma的出现密切反映了它们的进化起源。
简介
早期动物进化的时间是一个有争议的问题,观点通常分为两大学派。一种观点认为,化石记录相对准确,寒武纪辐射期间主要动物群体的出现密切反映了它们的进化起源1,2,3..相反的观点,主要基于分子钟估计的分歧日期,认为有更深刻的进化史4,5,6,7.近年来,这些观点之间的分歧已经缩小8,并确认埃迪卡拉生物群的某些成员可能是真后生动物9,10更精确的分子分化估计更接近化石记录6,11.然而,这些观点之间仍然存在不一致,特别是关于寒武纪Terreneuvian系列类群的经典“爆炸”(包括微量化石)是否代表了双侧冠群的真正辐射,或者这些冠群的起源是否更早地在埃迪卡拉纪被发现。
三叶虫是全群真节肢动物的一个分支,其首次出现基准(FAD)标志着Terreneuvian系列和临时寒武纪系列2(目前可追溯到约521 Ma)之间的边界。12,13.他们是最大和最成功的古生代群体之一,存在了大约2.7亿年,有超过22,000个已描述的物种14,15.这些优秀的化石记录——由于它们易于保存,生物矿化的外骨骼在生命中多次蜕皮——可以用来解决有关早期动物进化的重要问题16,17.三叶虫被认为是深两侧动物(因此是后生动物)分化年代论证的典范,并形成了这一观点早期支持者论证的重要组成部分4,18,19.其中一个原因是,三叶虫在化石记录中出现时,被认为表现出了大量的省性,在寒武纪早期被分为两个主要的生物地理区域:“olenelline”省(例如,劳伦蒂亚,波罗的海)和冈瓦纳的“redlichiine”省(包括南极洲,澳大利亚,中国和印度等地区),过渡区(有时被称为“bigotinid”省)出现在西冈瓦纳和西伯利亚等地区20.,21,22(无花果。1;这里我们使用子序23).一般认为,三叶虫必须有一个神秘的进化史才能产生这样的模式,而所观察到的分类单元分布是地理分隔的结果,在这种情况下是由于超大陆分裂和随后某些古大陆的隔离。这些模式要么与罗迪尼亚的分裂有关(约700-800 Ma)19或短暂的Pannotia(约550-600 Ma)24,25,26尽管分子钟估计的改进表明前者尤其不太可能。考虑到三叶虫作为全群真节肢动物的公认地位27这些生物地理模式与新元古代超大陆分裂的联系,已经被用来支持节肢动物进化的深刻、神秘历史,以及更普遍的早期动物进化的论点4,18,19,24,25.然而,这一推理是基于两个主要假设:(a)最早的三叶虫已经表现出确定的生物地理地方性和系统发育多样性;(b)观察到的生物地理模式来自于地理分隔而不是扩散。它还提出了关于早期三叶虫化石记录的问题,这些问题不容易回答。例如,如果生物矿化的外骨骼和相关特征是该群体的突触形态,为什么特勒叶三叶虫在化石记录中缺席,尽管在同一时期有足够的雪利记录?

颜色显示了所谓的“olenelline”(蓝色)和“redlichiine”(红色)早寒武纪三叶虫动物区系的大致区域,以及它们之间的过渡区域(紫色)。重要的是,这种地方性在最早的三叶虫动物群中并不明显,这些三叶虫动物群来自西伯利亚和西冈瓦纳的过渡地区,以及劳伦蒂亚(详情见正文)。注意,北(n)和南(s)的位置中国在寒武纪地理重建中一直不确定,并被标记为这样(?)。基于Torsvik & Cocks图2.7的地图92.
在这里,我们回顾了早期三叶虫进化的神秘历史的证据。我们还回顾了支持地理分隔模型背后的假设,鉴于>30年的工作极大地提高了我们对导致现代生物地理模式的因素的理解,包括地理分隔和扩散的相对重要性。
生物矿化问题
尽管有人提出了不同的观点28,29,三叶虫(可能不包括不可知论者30.)现在几乎被普遍接受,形成了一个由几种突触形态定义的分支,包括背侧外骨骼的钙化,以及具有钙化晶状体和环形缝合线的眼睛31,32,33.背面缝合线也可能是该组的突触形态,根据其在该组中的缺失分别代表祖先状态或继发损失,不包括或不包括奥内线(在eodiscids中,背面缝合线也可能独立衍生16).因此,任何生活在FAD之前(约521 Ma)的祖先三叶虫都应该拥有生物矿化的外骨骼(除非二次丢失);然而,没有发现Terreneuvian三叶虫,尽管在这一时期的大部分时间里存在着不同的shelly动物群。任何关于Terreneuvian(或前寒武纪)三叶虫因为没有矿化而在记录中缺失的观点都意味着所有主要的三叶虫谱系都必须在这一早期时期的某个时候分开,然后在大约521 Ma或此后不久独立地进化出钙质外骨骼。这一论点有效地抛弃了对进化支突触形态的定义,暗示三叶虫是目前所认为的多系——大多数工作者会拒绝这种观点。对于这种猖獗的趋同,已经有了争论,例如,海水化学的变化引发了这一时期几个双侧动物谱系中钙质骨骼的发展16.然而,上面提到的与生物矿化相关的特征在所有假设跨越这一边界的世系中独立发展仍然是极不可能的。例如,基于Paterson等人最近的寒武纪三叶虫系统发育分析。16这意味着11个独立的钙化外骨骼和相关的眼睛结构,7个背部面部缝合线(基于他们的多时代时钟模型,忽略521到522 Ma的差异,以考虑记录中的不确定性;看他们的图16).此外,如果按照目前的设想,三叶虫确实独立进化出了这些特征,这就意味着当时存在大量非矿化的三叶虫姐妹群,人们应该会在寒武纪早期看到其中一些Lagerstatten.然而,从这些沉积物中还没有可以被认为是“非矿化三叶虫”的例子(注意,任何这样的分类单元都必须与特定的三叶虫群比其他三叶虫更密切相关)。所有这些生物似乎都不太可能在大约521 Ma和这些沉积物中最古老的(例如,澄江生物群)之间突然灭绝,或者由于某种原因没有保存下来。
在没有任何其他令人信服的理由来怀疑三叶虫化石记录的准确性的情况下,因此,假设上面讨论的特征代表了真正的突触形态,并且三叶虫的出现接近它们的FAD是更加狭隘的。值得注意的是,尽管不可知者的位置对本文提出的论点没有太大影响,但从三叶虫中排除这一群体表明可能独立获得了钙化的背侧外骨骼。
Terreneuvian的节肢动物痕迹
微量化石也被用来证明三叶虫进化的神秘历史。节肢动物的踪迹来源于特勒努斯等Rusophycus被认为是三叶虫34(尽管出现在人类首次出现之前的1000 - 1500万年35),尽管一些作者后来提出这些可能是由其他节肢动物产生的36,37.三叶虫是寒武纪真性节肢动物的一个更大的多样性的成员,其中绝大多数是非矿化的。其中许多(如三叶虫)在前后轴上表现出一系列的双分枝(“双分枝”),带有颌基的附属物38,39,40.特别是,拟足纲(一种类似三叶虫的大型真节肢动物,包括三叶虫)通常有非常相似的附属物38,这显然是该群体的原始特征。许多洋蹄动物也表现出与三叶虫相似的整体形态,其中一些可能填补了类似的生态位。因此,可以预期,这样的分类群在剑齿虎的早期历史中产生了类似的痕迹,这一定发生在三叶虫的FAD之前(图2)。2).甚至有可能更多的基底非矿化茎类真节肢动物(例如,fuxianhuiids)41而且Parapeytoia42它们表现出相同的基本附属物结构)可能会产生类似的痕迹。寒武纪二系(阶段四)新近解释Rusophycus被认为是由一种非矿化甲壳类节肢动物产生的43支持了这些早期痕迹可能由非三叶虫产生的观点。也有人指出,虽然化石的痕迹很像Rusophycus而且爬迹发生在二叠纪大灭绝之后(例如,在三叠纪44,45),这并不被认为是后二叠纪三叶虫的证据46(这些也被归为甲壳类类群)。为什么我们要从不同的角度考虑这些痕迹在公元前521 Ma之前的存在,而其他明显的可能产生这些痕迹的候选者也存在?基于以上,对三叶虫化石记录的更字面的解读与微量化石记录并不不一致。相反,它支持对这些痕迹的解释,这些痕迹代表了从早期Terreneuvian开始的全群真节肢动物的早期多样化,更衍生的Terreneuvian类型的分类群(例如,更“典型的”Rusophycus发生在阶段235),并为三叶虫在大约521 Ma的FAD之前的进化提供了额外的时间。

这一类群的一个共同特征是它们相似的双栖附属物,很可能一些非三叶虫的偶蹄动物产生了与三叶虫相同的痕迹(例如,Rusophyscus,爬迹).如果三叶虫的出现时间接近它们在化石记录中出现的时间(约521 Ma),那么一定有大量的拟洋脚类(以及其他更遥远的类群)的早期历史,从而排除了认为三叶虫在Terreneuvian时期产生这些痕迹的要求。基于Ortega-Hernández等图6b的拓扑结构。38.
地理分隔假设
相关陆生和淡水类群被海洋盆地分离的分布可以用大陆分离导致的地理分隔或海洋扩散(即跨海洋的远距离扩散)来解释。在物种起源47达尔文认为,偶然的散布事件(虽然罕见,但在漫长的地质时期内经常发生)可以解释这种分布,并进行了各种实验,以证明某些植物种子在长时间浸泡在盐水中、或在作物和鸟类的消化道中仍能存活。他基本上反对只用地理位置划分的解释,他认为成功生物的扩散可能解释了化石动物群在相同的地质时间间隔内(即使相隔很远)是最相似的现象,而这一点得到了不同大陆的海洋动物群比陆地动物群更相似的支持47.达尔文还指出,火山岛屿上的生物一定是通过分散到达的,这在今天仍然是一个强有力的论点——在地质时间尺度上,这类事件在海洋岛屿上的重要性是不可否认的48,49.然而,大约从20世纪60年代开始,板块构造的验证和分支学的兴起提供了一个令人信服的由系统发育假说验证的大陆碎片化的地理分隔模型,而地理分隔(或分支学)生物地理学成为主导范式50.相比之下,海洋(或达尔文所说的“偶然”)扩散的假设被认为是不科学的,因为它们无法被证伪,基本上每一种模式都可以通过调用一系列扩散事件来解释50(尽管达尔文指出,洋流和盛行风等控制传播的因素并不是“偶然的”47).然而,这种对三叶虫生物地理模式的解释是在地理分隔必须负责的一般假设下出现的。
在过去25年左右的时间里,海洋扩散在解释现代生物地理分布方面发挥的作用比以前认为的要大得多,这一点已经变得越来越清楚50,51,52,而先验地排除扩散作为一个因素的分隔生物地理学方法受到了广泛的批评53,54,55.分子测年表明,许多被认为代表大陆分离的地理分隔实例的群体内的系统发育分裂发生得太迟了,以至于事实并非如此50.断层分布通常在一定程度上受到构造运动的控制,但最近的研究表明,替代信号经常或多或少地被海洋扩散的实例所覆盖,形成了一个复杂的模式网络,在不同的群体中各不相同56.即使对于曾经被认为表现出经典冈瓦纳地理分布(即与非洲、南极洲、澳大利亚、印度和南美洲的分裂有关)的群体,包括慈鲷和其他淡水鱼,也是如此57,58,以及各种植物群,包括南洋杉而且假山毛榉56,59.这并不意味着地理分隔在解释生物地理分布方面是一个无关紧要的因素,它仍然被认为是某些情况下的主导因素(例如在南半球陆地动物中)56).然而,现在很清楚的是,我们不能再简单地假设地理分隔是控制这种分布的主要因素。
早期的研究表明,地理分隔是解释寒武纪早期三叶虫生物地理模式的主要因素4,18,19指出,最早的三叶虫所表现出的省域性基本上证明了“化石记录中没有记录的较早阶段的地理分隔”(Fortey et al。19,第18页)。再加上这个推理中隐含的假设,即地理分隔一定是这种模式的原因(而不是扩散),早期分子钟提出的极深的差异日期表明,三叶虫可能有一个隐秘的历史,可以追溯到几亿年前,而与罗迪尼亚分裂有关的地理分隔可能是观察到的生物地理模式的原因。
最近,一些使用改良版布鲁克斯简约分析的研究表明,三叶虫的生物地理模式可能是由更近的、短暂的(也有争议的)三叶虫的分裂造成的60,61)新元古代晚期的超大陆Pannotia24,26这与最近对分子钟的估计一致6,11.这些方法包括用生物地理分布替换系统发育中的终端分类单元和内部节点,并将由此产生的信息编码在单独的字符矩阵中,其中“分类单元”代表生物地理区域62.然后对该矩阵进行简约分析,得出的树拓扑结构被解释为反映动物群连接的历史(在这种情况下,古大陆之间)。“地理分布”树反映了区域之间屏障的建立,而“地理分布”树反映了屏障的消除24,62.因此,重要的是要注意,地理扩散代表了与海洋扩散完全不同的概念。事实上,这些模式中没有任何机制允许真正的海洋扩散,因为解决的任何关系都被解释为由于在这些区域之间建立或消除障碍(例如与构造运动或海平面波动有关)而导致区域隔离或合并。因此,这样的模型可以产生很好解决的“地理位置”树,不一定是地理位置的结果。例如,受大陆间相对距离和洋流等因素影响的扩散,可以产生不同类群中相似的联系历史50,这是现代海洋动物分布的主要控制因素。如上所述,尽管系统发育关系显然与替代假说相一致,但在许多现代案例中,分子定年法支持对海洋扩散的解释50.很明显,当从分散和分离的角度解释生物地理模式时,分歧的时间是一个关键问题。最近的形态学时钟估计表明三叶虫最有可能出现在福图尼纪16(可能高估了;(见下文)代表了与这些现代案例类似的情况,该群体的年龄显然是在大陆分离的估计之后。就像这些现代的例子一样,如果由于构造事件和分类单元分化估计之间的时间不匹配,地理分隔解释被证明是不一致的,那么海洋扩散很可能是罪魁祸首。
分散:解释海洋无脊椎动物分布的一个主要因素
在解释现代生物地理分布时,关于地理分隔(通过大陆分离)与海洋扩散的重要性的争论可能在很大程度上与三叶虫的情况无关,因为这种形式的地理分隔通常只用于解释陆地和淡水类群的分布50.在现代海洋动物中——包括像大多数早期三叶虫那样局限于大陆架的分类群——地理分隔的实例几乎总是涉及到通过陆地(而不是像寒武纪三叶虫那样通过海路)来分裂种群。63,64,65.今天,海洋无脊椎动物的分布——包括被深海盆地(如大西洋、太平洋)隔开的动物群的差异——受到与一系列复杂因素相关的不同扩散能力的控制,这些因素包括洋流、物理屏障和纬度/温度梯度,而不是这些海道首次开放时的遗迹迁移模式66,67.因此,将大陆分离作为解释三叶虫分布的主要因素与已知的现代海洋动物群不一致。
在解释寒武纪三叶虫的生物地理模式时,快速扩散(从地质学角度来说)被认为不太可能,而是一段神秘的历史,需要大量聚集或罕见16.事实上,海洋无脊椎动物通常可以在很远的距离内迅速分散66无论是通过浮游幼虫的扩散,还是几乎可以肯定存在于寒武纪的其他措施,例如在半淹没底物上的运输,如漂浮的大藻类或浮石筏68,69.例如,即使在过去的几百年里,浮石筏也经常出现在所有主要的海洋中,并已被证明发展了不同的组合,并将生物远距离转移(例如,在最近的事件中,>有80种,>有5000公里70).南半球沿海群落也通过大型藻类筏频繁的长距离传播在生物上联系在一起69.这些筏子有助于确认海洋无脊椎动物的分布受到洋流的强烈控制70,71,72.重要的是,尽管这些载体的大多数招募是由具有远洋幼虫的类群进行的,但缺乏这些阶段的底栖海洋无脊椎动物也会定期通过浮石和大型藻类筏进行运输68,73,74.我们有理由假设三叶虫(尤其是小的个体)也可能是,而且在寒武纪,传播是海洋无脊椎动物分布的主要驱动力,就像今天一样。
在最早的三叶虫中缺乏省域性和系统发育多样性
有人反复指出,即使在它们最早的历史中,三叶虫也表现出明显的生物地理地方性和系统发育多样性4,16,18,19,20.,75被分为“olenelline”省和“redlichiine”省。如果三叶虫确实从它们在化石记录中出现的角度就表现出明显的地域性,那么这可能被认为是一段神秘历史的有力论据。然而,即使对数据进行简单的检查,也表明这一观点并没有得到很好的支持。
很明显,最早的三叶虫出现在劳伦蒂亚、西伯利亚和西冈瓦纳的同一时期76,77,最初包括来自龟甲科(Archaespididae)和龟甲科(fallotaspidoids)的olenelline,以及来自龟甲科(Bigotinidae)的redlichiines78,79,80,81,82(无花果。3),这两个物种被认为是密切相关的80.尽管存在地域主义的争论,但在所有这三个地区都存在类似的fallotaspidoids,而在Laurentia显然没有bigotinids。中国和东冈瓦纳地区最古老的三叶虫是原始的赤藓虫,比如Parabadiella,其年代有些争议76,77,83.在澳大利亚,最近有人提出,这些三叶虫与上述地区最古老的三叶虫几乎同时出现;84,85然而,这些东冈瓦纳三叶虫与在西冈瓦纳序列中发现的redlichioids有相似之处86.无论如何,这些都清楚地代表了可能与bigotinids密切相关的原始形式16,80.在波罗的海和阿瓦洛尼亚,最早出现的三叶虫是衍生的olenelline holmiids,通常被认为比上面讨论的分类群要年轻一些76.

一个红叶三叶虫目的高级分类学,主要在Adrain目之后23.请注意,这并不代表所示的各个类群中的分类单元的详尽列表,仅代表文中讨论或中所示的类群b.b早寒武纪三叶虫的系统发育(修改自Paterson等的图2)。16;有关完整的物种名称和年代参考资料,请参阅本出版物)。尖端代表了所包含物种的年龄,但是尽管我们试图保持近似的相对内部分支长度(同时向树的底部增加速率),这些并不是精确的比例。该树位于Olenellina亚目和Redlichiina亚目之间(见正文),分化日期为521.5 Ma,以说明相对准确的化石记录情景。早期红褐石属的比较Bigotina bivallata(c,d)和早期olenelline fallotaspidoidProfallotaspis tyusserica(e,f).这些类群表现出相当大的形态相似性,支持这些类群之间的密切关系。经Bushuev等人许可修改。78和盖尔;80比例尺1毫米。
基于此,很明显,在三叶虫最早出现的地区,出现在最低的几个三叶虫生物带内的动物群(可能代表了几百万年)是相似的,并且包含或多或少密切相关的原始形式,这与最早的三叶虫“确定的省性”的主张直接矛盾。只有在这段时间之后,两个主要生物地理省份相对不同的动物群才得以建立,基本上涉及到冈瓦纳(和其他地方)的redlichiines的辐射,以及劳伦西亚的olenellines的持续主导地位。然而,这些高级分类群有明显的重叠,缝合三叶虫迅速出现在所有地区。以前有人认为,这种模式可能是由于寒武纪早期(新元古代分裂之后)持续的裂谷作用造成的,最初古大陆之间的分散更容易,后来随着大陆分离的继续,动物群变得更加局限87.这说明了分布如何可以反映构造关系,而不需要严格的大陆破碎度分隔。寒武纪早期世界范围内的三叶虫生物地理模式实际上还没有被非常详细地探索(尽管参见Álvaro等。75),应进行更彻底的调查。例如,上面讨论的“不同”动物群中元素的重叠表明,这种模式可能由更连续的纵向梯度更好地解释,从西部的劳伦蒂亚到东部的冈瓦纳(图2)。1).
最早的三叶虫显示出系统发育多样性的说法似乎也没有什么根据。如上所述,最早出现的三叶虫是olenelline fallotaspidoids和redlichiine bigotinids,尽管这些类群在很大程度上是根据背部面部缝合线的存在(Redlichiina)或不存在(Olenellina)而分为不同的亚目,但这些类群在形态上表现出明显的相似性。例如,一个比较的bigotinidBigotina bivallata(无花果。3 c, d)和fallotaspidoidProfallotaspis tyusserica(无花果。3 e, f)显示出相当的整体比例,眼脊的形状和方向,以及这些眼脊如何从眉间前部延伸——这似乎是最早的三叶虫的原始特征80.一般认为,早期缺乏背侧面部缝合线的olenellines(如fallotaspidoids)代表祖先的状态。我们一致认为这是最节俭的(虽然不是唯一的)选择,并且在面前线的存在p . tyusserica(无花果。3 e, f)可能为三叶虫背部缝合线最初是如何进化的提供了线索78.虽然这一物种的面部后缝合线似乎没有,但前缝合线(连接环缝和吻侧/边缘缝合线)的打开会大大扩大蜕皮口,也会使“自由”脸颊在蜕皮过程中更容易脱落,就像在这里重新设计的标本中所发生的那样(图2)。3 e, f).面部前缝的存在p . tyusserica,以及非常相似的整体形式的本种和bigotinids如b . bivallata,表明这些形态可能接近于缝合的红苔藓和未缝合的olenellines之间的分裂,这两个类群的最早代表基本上通过这些形式表现出连续的变化。因此,将最早的赤藓属和olenellines分为不同的亚目,主要是基于背侧面部缝合线的存在/不存在,这大大夸大了最早的三叶虫中存在的形态变异的数量。这说明了分类系统在靠近其起源的地方划分高级分类群时不可避免的局限性(我们必须在某个地方“划清界限”)。
时间和形态空间的多样化
对三叶虫化石记录进行更字面上解释的一个重要考虑是,该模型与假设的系统发育关系和观察到的随时间变化的多样性模式的一致性。数字3 b显示了最近寒武纪三叶虫系统发育的简化版本(基于Paterson等人的“放松时钟”模型)。16),其(修正的)发散日期为521.5 ma,说明了相对准确的化石记录的情景。在这个假设下,并考虑到可能原始的背面缝合线的缺失,在Fallotaspididae上生根的原始树是合理的。然而,考虑到最古老的fallotaspidoids(不包括在分析中,例如,Profallotaspis)倾向于表现出与bigotinids更中间的形态,可以提出一个论点,即将树的根更靠近这些组之间的分裂,这是图中所示的。3 b.这对这里提出的论点没有什么不同,尽管它确实说明了奥莱奈利纳和雷德利希纳之间很早就有了分歧,这可能是基于化石记录的预期。在任何一种情况下,Fallotaspidoidea都被认为是para aphytic(树底部的深色分支),从而产生了一个多系的Olenelloidea。这支持了fallotaspidoid型形态是一种原始特征的想法,存在于树的底部。它还支持了原始redlichioid(例如,bigotinids, abadiellids)从fallotaspidoid型形态(我们注意到在“epoch clock”模型下)的早期分化16这些建议是稍微派生的)。
如果三叶虫有大量的神秘历史,我们可能会看到不同的形态在时间和形态空间上随机出现,就像以前看不见的谱系在化石记录中出现一样。然而,总的来说,地层的外观似乎密切地反映了系统发育,这表明情况并非如此。最初,我们看到了非常少量的相关科,随后出现了更多派生的类群,这被解释为从这些古老的分类单元中多样化。例如,在形态上有一个明显的梯度,从法罗草皮类,到二丁酸类80和早期的红鲤类Lemdadella88而且Parabadiella83,到更年轻,更衍生的形式,如Eoredlichia89而且Redlichia39.同样的情况也适用于奥烯类:从早期的fallotaspidoids,到后来的例子,如Nevadia而且Judomia,到更年轻的奥烯类奥氏虫而且钬氧化物82(无花果。3 b).其他早期类群,如椭圆头类,很可能是从早期的redlichioid或bigotinid型形态进化而来的(Bigotinidae有时也包括在这个类群中23).随着时间的推移,这种外观和多样化的模式表明,我们观察到的是真实的辐射,没有必要引用一个扩展的神秘进化史来解释寒武纪早期三叶虫的生物地理分布或系统发育关系。
过去的推动
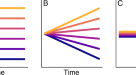
最近的研究90,91他们强调,从统计上看,存活较长时间的成功演化支可能具有较高的初始多样化率,这种效应被称为过去的推动(POTPa)。与初始多样化率较低的演化支相比,这样的演化支在任何给定的时间长度内都有更高的生存机会。POTPa的规模由人员流动率控制;即,对于给定的多样化速率(物种灭绝速率),更高的POTPa意味着高的灭绝速率。这种效应很可能会显著缩短最近形态时钟所预测的神秘历史16这表明三叶虫很可能出现在福图尼纪。这些作者指出,他们的预测必然是基于树木可见部分的比率(在519 Ma之后,他们的分析中包括了最古老的三叶虫的年龄),如果在此之前比率更高,那么估计的分歧日期将被高估16.他们还表明,强制执行522 Ma的分歧日期将需要在树的底部提高比率(正如POTPa所预期的那样),但这对他们的整体结果几乎没有影响;整个寒武纪的速率保持相对均匀16.这表明,任何对背景速率的恢复都发生得很快,他们在寒武纪早期观察到的略高的速率可能是由于接近三叶虫出现时的短期快速多样化造成的。因此,我们认为他们的分歧估计之间的不匹配(在误差条内,年轻至约526 Ma)和更字面的解读三叶虫化石记录可能是由影响,如POTPa(图。4).还应该注意的是,POTPa表明,Terreneuvian中三叶虫的缺失不太可能用稀有或低多样性来解释,因为这样的群体极有可能长期存在而不灭绝90,91.事实上,POTPa预测物种多样性最初会迅速增加,这意味着像三叶虫这样具有很高保存潜力的化石群应该在出现后不久就会出现在记录中。

蓝色实线显示了一个具有大量POTPa的场景,这可能是对像三叶虫这样极其多样化和成功的进化支的预期。在下降到背景率之前,多样化率(线的斜率)最初非常高。红色虚线显示了三叶虫多样性在该类群未观察到的早期历史中的投影,基于树的“可见”部分——在这种情况下是在519 Ma之后(由垂直虚线显示),类似于Paterson等人对寒武纪三叶虫系统发育和形态时钟分歧的估计。16.像POTPa这样的生存偏差可能解释了这种估计与三叶虫化石记录之间的差异。
结论
Terreneuvian或更早的非矿化三叶虫的建议被证明是非常不狭隘的,这意味着在所有主要的早期三叶虫谱系中,与外骨骼生物矿化相关的结构普遍收敛,并且放弃了联合分支的突触形态。这表明,没有可信的理由提出没有Terreneuvian三叶虫化石记录,假设一个实质性的神秘的进化史。尽管先前的陈述与此相反,当三叶虫出现在化石记录中时,它们表现出有限的地域性和相对较低的系统发育多样性。即使在寒武纪系列2的其余部分中出现了更多不同的动物群,这些动物群之间也有相当大的重叠,多样化的模式表明这是实时发生的(而不是在三叶虫FAD之前的分化所导致的)。鉴于过去几十年里我们对地理分隔和传播在解释现代生物地理模式方面的相对重要性的理解发生了变化,而且普遍观察到现代海洋无脊椎动物群不表现出由大陆分离引起的地理分隔模式,三叶虫的生物地理模式不太可能是由这种形式的地理分隔造成的。最近对形态时钟的估计(表明三叶虫可能出现在福图尼纪)和对化石记录的更字面的解读之间的不匹配,可以用诸如过去的推动等效应来解释,过去的推动预示着在演化支的初始辐射期间会有更高的多样化率——尤其是在像三叶虫这样非常长寿和成功的群体中。我们的结论是,三叶虫的FAD紧密地反映了它们的进化起源,并且没有令人信服的证据表明这一群体有一个扩展的神秘进化史。
报告总结
实验设计的进一步信息可在自然研究报告摘要链接到这篇文章。
参考文献
寒武纪化石记录和门的起源。中国。广告样稿,杂志。43, 157-165(2003)。
Budd, g.e. & Jensen, S.对双侧动物门化石记录的批判性重新评价。医学杂志。牧师。75, 253-295(2000)。
布德,g.e.,曼恩,r.p.茎和冠群的动态。科学。睡觉。6, eaaz1626(2020)。
库珀,A. &福泰,R.进化爆炸与系统发育融合。生态发展趋势。另一个星球。13, 151-156(1998)。
动物身体计划的起源:从化石证据和调控基因组的观点。发展147, dev182899(2020)。
欧文,D. H.等人。寒武纪难题:动物早期历史上的早期分化和后来的生态成功。科学334, 1091-1097(2011)。
斯珀林,E. A.和斯托基,R. G.早期动物进化的时间和环境背景:考虑“爆炸”的所有成分。中国。广告样稿,杂志。58, 605-622(2018)。
坎宁安,刘安根,本特森,S. &多诺霍,p.c.j .动物起源:分子钟和化石记录能协调一致吗?BioEssays39, e201600120(2017)。
邓福生,刘安国,唐诺修,刘志刚。埃迪卡拉纪发育生物学。医学杂志。牧师。93, 914-932(2018)。
埃文斯,S. D.,休斯,I. V., Gehling, J. G. & Droser, M. L.南澳大利亚埃迪卡拉纪最古老的双侧体化石的发现。国家科学院学报美国117, 7845-7850(2020)。
李,M. S. Y., Soubrier, J. & Edgecombe, G. D.寒武纪大爆发期间表型和基因组进化的速率。咕咕叫。医学杂志。23, 1889-1895(2013)。
马鲁夫,a.c.等。最早的寒武纪动物和海洋地球化学变化记录。青烟。l Soc。点。公牛。122, 1731-1774(2010)。
马鲁夫,a.c.等。摩洛哥Nemakit-Daldynian-Tommotian边界δ13C位移持续时间对早寒武纪碳循环的制约。地质38, 623-626(2010)。
奥陶系三叶虫分布与多样性概要。在“早期古生代生物地理学和古地理学”(ed . A. T. Harper, T. Servais)。青烟。Soc。Lond。Mem.38, 297-336(2013)。
三叶虫的问题:分类、系统发育和隐生问题。青烟。玛格。157, 35-46(2020)。
帕特森,J. R., Edgecombe, G. D. & Lee, M. S. Y.三叶虫进化速率限制寒武纪大爆发的持续时间。国家科学院学报美国116, 4394-4399(2019)。
霍姆斯,J. D.帕特森,J. R. & García-Bellido, D. C.南澳大利亚寒武纪早期三叶虫的复杂轴向生长模式。Proc. R. Soc。B。288, 20212131(2021)。
福泰,R. A.布里格斯,D. E. G. &威尔斯,M. A.寒武纪进化“大爆发”重新校准。Bioessays19, 429-434(1997)。
福泰,R. A.,布里格斯,D. E. G. &威尔斯,M. A.寒武纪进化的“爆炸”:从形态差异中解耦分叉。医学杂志。j·林恩。Soc。朗。57, 13-33(1996)。
福泰,r.a. &欧文斯,r.m.。主要进化辐射139-164(克拉伦登出版社,1990)。
皮罗拉,G. L.三叶虫杜Cambrien inférieur杜西南德拉萨代涅,意大利。Palaeontogr。斜体字。78, 1-173(1991)。
早寒武纪的三个动物区系。Proc。日本。专科学校48, 242-247(1972)。
德莱恩,j.m.三叶虫瓦尔奇,1771年。据研究3148, 104-109(2011)。
利伯曼,B. S.测量寒武纪辐射的脉冲。中国。广告样稿,杂志。43, 229-237(2003)。
一些基本的早寒武纪三叶虫的系统发育分析,Eutrilobita的生物地理起源,以及寒武纪辐射的时间。j . Paleontol。76, 692(2002)。
Meert, J. G. & Lieberman, B. S.关于最新新元古代和早寒武纪构造事件的古地磁和古生物地理学观点。j .青烟。Soc。Lond。161, 477-487(2004)。
休斯,北卡罗来纳州。三叶虫身体图案的进化。为基础。地球行星。科学。35, 401-434(2007)。
Lauterbach K.-E。Schlüsselereignisse在der Evolution des Grundplans under Arachnata(节肢动物)。Abh。Verh。Naturwiss。版本。Hambg。23, 163-327(1980)。
Lauterbach K.-E。三叶虫和蛛形纲。黑旋风。澳新银行。210213-238(1983)。
莫伊修克,J.- b .卡隆。伯吉斯页岩化石揭示了不可知论者的问题。Proc. R. Soc。B。286, 20182314(2019)。
Edgecombe, g.d. & Ramsköld, L.寒武纪蛛形目的关系与三叶虫的系统位置。j . Paleontol。73, 263-287(1999)。
Ramsköld, L. & Edgecombe, G. D.三叶虫单系重访。嘘。医学杂志.4, 267-283(1991)。
福泰,R. a . &惠廷顿,H. B.三叶虫作为一个自然类群。嘘。医学杂志。2, 125-138(1989)。
前寒武纪晚期和早寒武纪地层的化石和对比。青烟。玛格。124, 97-119(1987)。
巴德,g.e.和杰克逊,i.s.c.寒武纪的生态创新和冠群门的起源。费罗斯。反式。r . Soc。B371, 20150287(2016)。
鲍德,郭志刚,李志刚。节肢动物的起源与进化。自然457, 812-817(2009)。
斯坦,M.,巴德,G. E.,皮尔,J. S. &哈珀,D. A. T.。Arthroaspisn. gen.是Sirius Passet Lagerstatte(寒武纪,北格陵兰)的常见元素,揭示了三叶虫的祖先。BMC另一个星球。医学杂志。13, 99(2013)。
Ortega-Hernández, J., Legg, D. A. & Braddy, S. J.无鳞节肢动物的系统发育及拟洋足纲的内部关系。支序分类学29, 15-45(2013)。
霍姆斯,j.d.,帕特森,j.r. & García-Bellido, d.c.三叶虫Redlichia来自下寒武统鸸鹋湾页岩Konservat-Lagerstatte南澳大利亚:分类学,个体发育学和软部解剖学。j .系统。Palaeontol。18, 295-334(2020)。
比克内尔,r.d.等人。寒武系真肢动物的咀嚼和硬噬作用的生物力学分析。Proc. R. Soc。B。288, 20202075(2021)。
杨,J.等。中国早寒武世富仙汇类揭示了真肢动物中颚基原足类的起源。Commun Nat。9, 470(2018)。
真节肢动物唇瓣的起源和进化。节肢动物结构。Dev。62, 101048(2021)。
普拉特,B. R.下寒武纪Rusophycus产于加拿大北极埃尔斯米尔岛:一种掠食性、非三叶虫节肢动物的化石。PALAIOS37, 165-184(2022)。
Zonneveld, j。,Pemberton, S. G., Saunders, T. D. A. & Pickerill, R. K. Large, robust Cruziana from the Middle Triassic of Northeastern British Columbia: Ethologic, biostratigraphic, and paleobiologic significance.PALAIOS17, 435-448(2002)。
布罗姆利,R. & Asgaard, U.格陵兰岛东部嘉士伯峡湾的三叠纪淡水ichnocoenses。Palaeogeogr。Palaeoclimatol。Palaeoecol。28, 39-80(1979)。
多诺万,S. K. Cruziana和Rusophycus:在某些情况下,三叶虫产生的痕迹化石?Lethaia43, 283-284(2010)。
达尔文,C。论自然选择的物种起源(约翰·默里,1859)。
Quammen, D。渡渡鸟之歌:灭绝时代的岛屿生物地理(哈钦森,1996)。
散布是海洋岛屿生物地理学和生物多样性演化的基础。j . Biogeogr。33, 193-198(2006)。
历史生物地理学中海洋扩散的复活。生态发展趋势。另一个星球。20., 68-73(2005)。
吉莱斯皮,R. G.等。长距离传播:假设检验的框架。生态发展趋势。另一个星球。27, 47-56(2012)。
德奎罗斯,A。《猴子的航行:不可思议的旅程如何塑造了生命的历史》(Basic Books, 2014)。
沃特斯,J. M.等。偏离轨道的生物地理学。系统。医学杂志。62, 494-498(2013)。
历史生物地理学中的模式选择揭示了在岛屿演化支中,始祖事件物种形成是一个至关重要的过程。系统。医学杂志。63, 951-970(2014)。
《泛生物地理学:起源、变形与衰落》。拉斯。比尔。33, 273-277(2007)。
Sanmartín, I. & Ronquist, F.南半球生物地理学推断基于事件的模型:植物与动物模式。系统。医学杂志。53, 216-243(2004)。
慈鱼科鱼类的地理分隔或跨大西洋扩散:分子证据的回顾。Hydrobiologia832, 9-37(2019)。
Capobianco, A. & Friedman, M.南半球淡水鱼分支的地理分布和扩散:古生物学视角:淡水鱼的地理分布和扩散。医学杂志。牧师。94, 662-699(2019)。
Noben, S.等人。冈瓦纳树蕨科的生物地理——地理分隔、扩散和灭绝的故事。j . Biogeogr。44, 2648-2659(2017)。
墨菲,J. B.等。Pannotia:为它的存在和地球动力学意义辩护。青烟。Soc。出版规范。503, 13-39(2021)。
埃文斯,检察官潘诺提亚被起诉。青烟。Soc。出版规范。503, 63-81(2021)。
李伯曼,B. S. & Eldredge, N.中泥盆世三叶虫生物地理学:地质过程和分析方法。古生物学22, 66-79(1996)。
Laakkonen, H. M., Hardman, M., Strelkov, P. & Väinölä, R.跨北极扩散和地理分隔的周期,以及amphi‐北方海洋动物的多样化。j .另一个星球。医学杂志。34, 73-96(2021)。
Bernardi, G, Findley, L.和Rocha-Olivares, A.在分离的海洋鱼类种群中,下加利福尼亚的地理分隔和扩散。进化57, 1599-1609(2003)。
米ams, A. G. K, Treml, E. A. Shields, J. L., Liggins, L. & Riginos, C.跨间歇性屏障的地理分隔和扩散:跨托雷斯海峡陆桥海洋动物的种群遗传结构。珊瑚礁30., 937-949(2011)。
遗传分化、生殖隔离与海洋物种形成。为基础。启生态。系统。25, 547-572(1994)。
Rosenblatt, R. H. & Waples, R. S.太平洋东部和中部海岸鱼类物种迁移种群的遗传比较:扩散还是地理隔离?Copeia1986, 275-284(1986)。
没有远洋幼虫期的海洋生物的扩散。中国。广告样稿,杂志。52, 447-457(2012)。
弗雷泽,c.i.等。南半球海岸通过频繁的长距离漂流活动在生物上联系在一起。咕咕叫。医学杂志。32, 3154 - 3160。e3(2022)。
布莱恩,S. E.等。通过浮石漂流快速、远距离地散布。《公共科学图书馆•综合》7, e40583(2012)。
布莱恩,S. E.等。2001-2002年西南太平洋浮石漂流和动物群分散:来自汤加的英安岩海底爆炸喷发的记录。地球的星球。科学。列托人。227, 135-154(2004)。
Jokiel, P. L. & Cox, E. F.在圣诞岛和夏威夷漂移浮石:海洋扩散模式的证据。3月青烟。202, 121-133(2003)。
弗雷泽,c.i.,尼库拉,R. & Waters, J. M.海洋漂流由沿海社区。Proc. R. Soc。B。278, 649-655(2011)。
尼库拉,R.斯宾塞,H. G.和沃特斯,J. M.被动漂流是跨洋基因流的强大驱动力。医学杂志。列托人。9, 20120821(2013)。
Álvaro, J. J.等。全球寒武纪三叶虫古地理的地方性简约分析。在“早期古生代生物地理学和古地理学”(ed . A. T. Harper, T. Servais)。青烟。Soc。Lond。Mem.38, 273-296(2013)。
Landing, E., Geyer, G., Brasier, m.d. & Bowring, s.a .寒武纪演化辐射:背景、相关性和年代地层-克服首次出现资料(FAD)概念的不足。Earth-Sci。牧师。123, 133-172(2013)。
张欣,等。寒武纪系列2和阶段3基础界定的挑战。Earth-Sci。牧师。172, 124-139(2017)。
Bushuev, E., Goryaeva, I. & Pereladov, V.最古老三叶虫的新发现Profallotaspis而且Nevadella在俄罗斯西伯利亚地台的东北部公牛。Geosci.89, 347-364(2014)。
内华达西部和加利福尼亚东部寒武纪第3阶段的岩石地层和生物地层。亩。北方。亚利桑那州。公牛。67, 26-42(2011)。
Geyer, G.已知最早的西冈瓦纳三叶虫来自摩洛哥的反阿特拉斯(Anti-Atlas),与Family Bigotinidae的修订版Hupé, 1953。自由/开源软件。stratocaster电吉他。64, 55-153(2019)。
摩洛哥fallotaspidid三叶虫重现。Beringeria18, 89-199(1996)。
帕尔默,a. R. &雷皮纳,L. N.透过黑暗的玻璃:小圆齿藻的分类、系统发育和生物地层学。大学,堪萨斯州。Paleontol。普通发布版。3., 1-35(1993)。
本特森,S.,康威·莫里斯,S.,库珀,B. J., Jell, P. A. & Runnegar, B. N.南澳大利亚的早期寒武纪化石。Mem。Assoc。Australas。Palaeontol。9, 1-364(1990)。
贝茨,M. J.等。南澳大利亚早寒武世年代地层学和地质年代学。Earth-Sci。牧师。185, 498-543(2018)。
贝茨,M. J.等。南澳大利亚早寒武纪的全球对比:雪梨动物群Dailyatia odyssei区。冈瓦那大陆Res。46, 240-279(2017)。
Hupé, P.贡献à l ' étude du Cambrien inférieur et du Précambrien III de l ' anti - atlas Marocain。德国矿业工业方向,德国矿业分类Géologie,服务器Géologique,注释等Mémoires103, 1-402(1953)。
布德,G. E. &詹森,S.化石记录的局限性和双侧螈起源的年代。系统。Assoc。出版规范。66, 170-193(2003)。
帕尔默,A. R. &罗威尔,A. J.寒武纪早期三叶虫,来自横贯南极山脉中部沙克尔顿石灰岩。Paleontol。Soc。Mem。45, 1-28(1995)。
戴涛,张欣。红叶三叶虫的个体发育Eoredlichia intermediata澄江Lagerstätte,下寒武纪,中国西南部。Lethaia46, 262-273(2013)。
巴德,g.e.和曼恩,r.p.早期动物进化中的生存和选择偏差和分子钟系统高估的来源。接口的焦点。10, 20190110(2020)。
巴德,g.e.和曼恩,r.p.历史是由胜利者书写的:过去的推动对化石记录的影响。进化72, 2276-2291(2018)。
托斯维克,T. H. &科克斯,L. R. M.早期古生代的新全球古地理重建及其世代。在“早期古生代生物地理学和古地理学”(ed . A. T. Harper, T. Servais)。青烟。Soc。Lond。Mem.38, 5-24(2013)。
确认
我们感谢Nigel Hughes和Greg Edgecombe的有用评论,极大地改进了手稿。我们也感谢Marissa Betts对这个工作的早期版本的评论,以及Ben Slater有深刻见解的讨论。
资金
乌普萨拉大学提供的开放获取资金。
作者信息
作者及隶属关系
贡献
J.D.H.构思了这项研究,并根据G.E.B.的意见撰写了手稿
相应的作者
道德声明
相互竞争的利益
作者声明没有利益竞争。
同行评审
同行评审信息
通信生物学感谢Nigel Hughes和Gregory Edgecombe对本工作的同行评审所作的贡献。首席处理编辑:Luke R. Grinham。同行评审报告是可用的。
额外的信息
出版商的注意施普林格自然对出版的地图和机构从属关系中的管辖权主张保持中立。
补充信息
权利和权限
开放获取本文遵循知识共享署名4.0国际许可协议(Creative Commons Attribution 4.0 International License),允许以任何媒介或格式使用、分享、改编、分发和复制,只要您对原作者和来源给予适当的署名,提供知识共享许可协议的链接,并注明是否有更改。本文中的图像或其他第三方材料包含在文章的创作共用许可中,除非在材料的信用额度中另有说明。如果内容未包含在文章的创作共用许可协议中,并且您的预期使用不被法定法规所允许或超出了允许的使用范围,您将需要直接获得版权所有者的许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/by/4.0/.
关于本文

引用本文
霍姆斯,j.d.,巴德,G.E.。重新评估早期三叶虫进化的神秘历史。Commun杂志5, 1177(2022)。https://doi.org/10.1038/s42003-022-04146-6
收到了:
接受:
发表:
DOI:https://doi.org/10.1038/s42003-022-04146-6
